

Divergent cytokine and transcriptional signatures control functional T follicular helper cell heterogeneity
- PMID: 40926076
- PMCID: PMC12479364
- DOI: 10.1038/s41590-025-02258-9
Divergent cytokine and transcriptional signatures control functional T follicular helper cell heterogeneity
Abstract
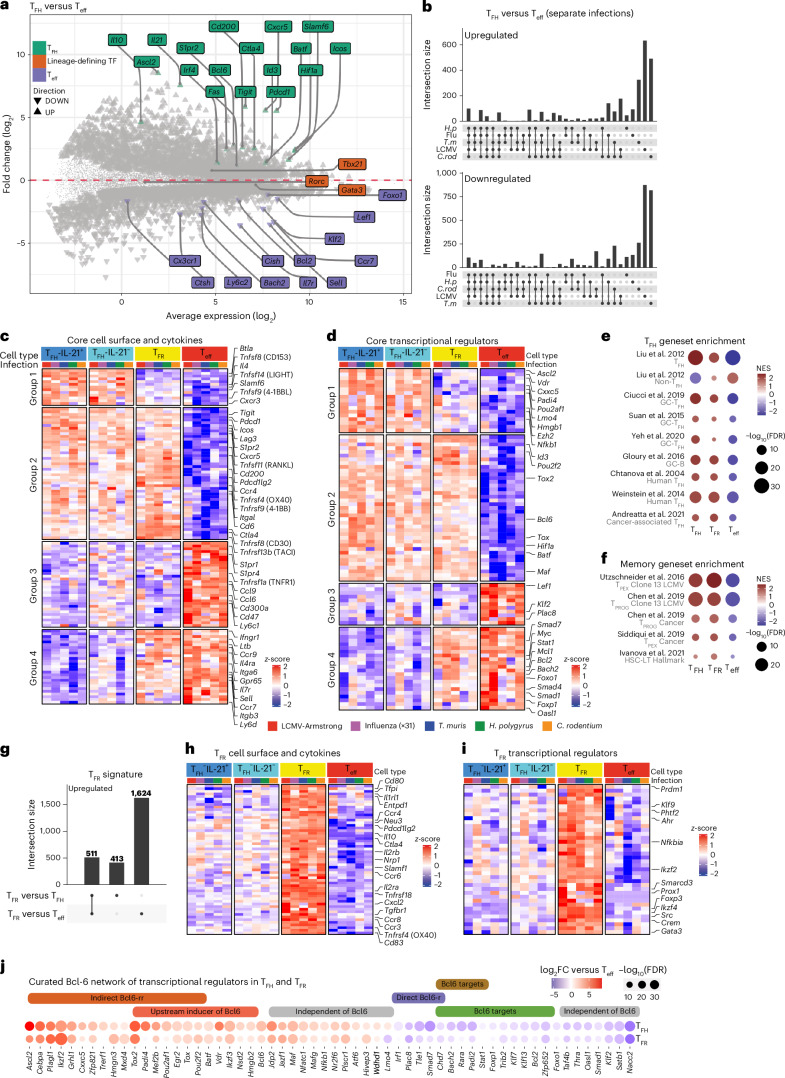
CD4+ T follicular helper (TFH) cells support tailored B cell responses against multiple classes of pathogens. To reveal how diverse TFH phenotypes are established, we profiled mouse TFH cells in response to viral, helminth and bacterial infection. We identified a core TFH signature that is distinct from CD4+ T follicular regulatory and effector cells and identified pathogen-specific transcriptional modules that shape TFH function. Cytokine-transcriptional TFH programming demonstrated that type I interferon and TGFβ signaling direct individual TFH phenotypes to instruct B cell output. Cytokine-directed TFH transcriptional phenotypes are shared within human germinal centers, but distinct TFH phenotypes dominate between donors and following immune challenge or in antibody-mediated disease. Finally, we identified new cell surface markers that align with distinct TFH phenotypes. Thus, we provide a comprehensive resource of TFH diversity in humans and mice to enable immune monitoring during infection and disease and to inform the development of context-specific vaccines.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: No authors declare competing interests.
Figures
















References
MeSH terms
Substances
Grants and funding
- GNT1137989/Department of Health | National Health and Medical Research Council (NHMRC)
- GNT2007812/Department of Health | National Health and Medical Research Council (NHMRC)
- GNT2007884/Department of Health | National Health and Medical Research Council (NHMRC)
- GNT1155342/Department of Health | National Health and Medical Research Council (NHMRC)
- GNT2033037/Department of Health | National Health and Medical Research Council (NHMRC)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
