

Distinct evolutionary trajectories of subgenomic centromeres in polyploid wheat
- PMID: 40926249
- PMCID: PMC12418699
- DOI: 10.1186/s13059-025-03759-4
Distinct evolutionary trajectories of subgenomic centromeres in polyploid wheat
Abstract
Background: Centromeres are crucial for precise chromosome segregation and maintaining genome stability during cell division. However, their evolutionary dynamics, particularly in polyploid organisms with complex genomic architectures, remain largely enigmatic. Allopolyploid wheat, with its well-defined hierarchical ploidy series and recent polyploidization history, serves as an excellent model to explore centromere evolution.
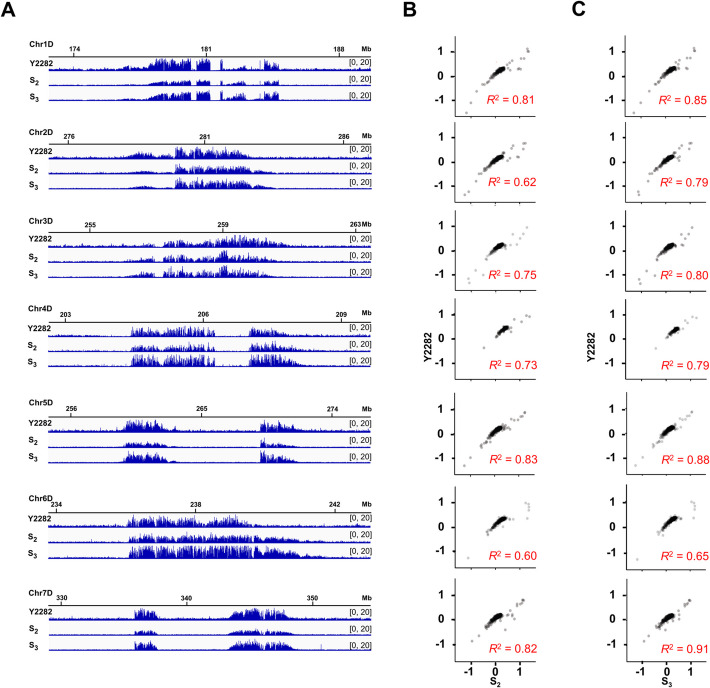
Results: In this study, we perform a systematic comparative analysis of centromeres in common wheat and its corresponding ancestral species, utilizing the latest comprehensive reference genome assembly available. Our findings reveal that wheat centromeres predominantly consist of five types of centromeric-specific retrotransposon elements (CRWs), with CRW1 and CRW2 being the most prevalent. We identify distinct evolutionary trajectories in the functional centromeres of each subgenome, characterized by variations in copy number, insertion age, and CRW composition. By utilizing CENH3-ChIP data across various ploidy levels, we uncover a series of CRW invasion events that have shaped the evolution of AA subgenome centromeres. Conversely, the evolutionary process of the DD subgenome centromeres involves their expansion from diploid to hexaploid wheat, facilitating adaptation to a larger genomic context. Integration of complete einkorn centromere assemblies and Aegilops tauschii pan-genomes further revealed subgenome-specific centromere evolutionary trajectories. By inclusion of synthetic hexaploid from S2-S3 generations, alongside 2x/6 × natural accessions, we demonstrate that DD subgenome centromere expansion represents a gradual evolutionary process rather than an immediate response to polyploidization.
Conclusions: Our study provides a comprehensive landscape of centromere adaptation, evolution, and maturation, along with insights into how retrotransposon invasions drive centromere evolution in polyploid wheat.
Keywords: Triticum aestivum; CENH3; CRW; Centromere evolution; FlLTR-RTs.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Competing interests: The authors declare no competing interests. Fangpu Han is a guest editor of the Centromere Structure and Evolution article collection, but was not involved in the editorial decision making or peer review of this manuscript.
Figures






References
-
- Lev-Yadun S, Gopher A, Abbo S. Archaeology. The cradle of agriculture. Science. 2000;288:1602–3. - PubMed
-
- Marcussen T, Sandve SR, Heier L, Spannagl M, Pfeifer M, Jakobsen KS, et al. Ancient hybridizations among the ancestral genomes of bread wheat. Science. 2014;345:1250092. - PubMed
-
- Avni R, Nave M, Barad O, Baruch K, Twardziok SO, Gundlach H, et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science. 2017;357:93–7. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
