

Proximity labeling of axonemal protein CFAP91 identifies EFCAB5 that regulates sperm motility
- PMID: 40931011
- PMCID: PMC12423330
- DOI: 10.1038/s41467-025-63705-7
Proximity labeling of axonemal protein CFAP91 identifies EFCAB5 that regulates sperm motility
Abstract
Radial spokes (RSs) are conserved multimolecular structures attached to the axonemal microtubule doublets and are essential for the motility control of both cilia and sperm flagella. CFAP91, an RS3 protein, is implicated in human male infertility, yet its molecular function remains poorly understood. Here, we demonstrate that Cfap91 knockout (KO) mice exhibit impaired sperm flagellum formation and male infertility. Using a transgenic rescue model expressing FLAG- and BioID2-tagged CFAP91, we reveal that CFAP91 immunoprecipitates with RS3 proteins CFAP251 and LRRC23, whose localization is disrupted in Cfap91 KO sperm flagella. In addition, proximity labeling in mature spermatozoa identifies EFCAB5 as a sperm-specific CFAP91-proximal component. We show that Efcab5 KO males exhibit reduced sperm motility and fertility. Our findings establish CFAP91 as an essential scaffolder of RS3 assembly and EFCAB5 as a sperm-specialized movement regulator, advancing understanding of axonemal specialization in mammalian spermatozoa and its relevance to male infertility.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

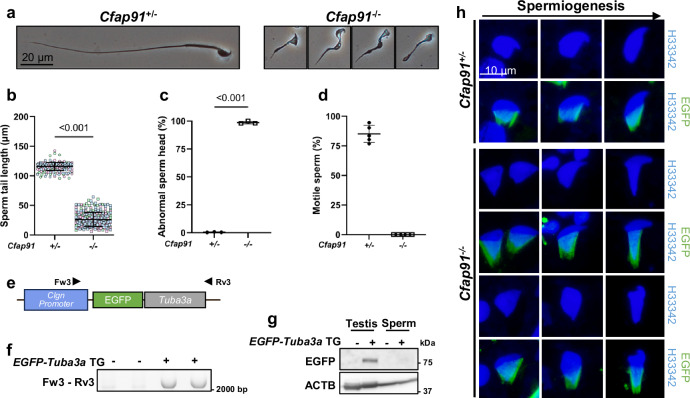
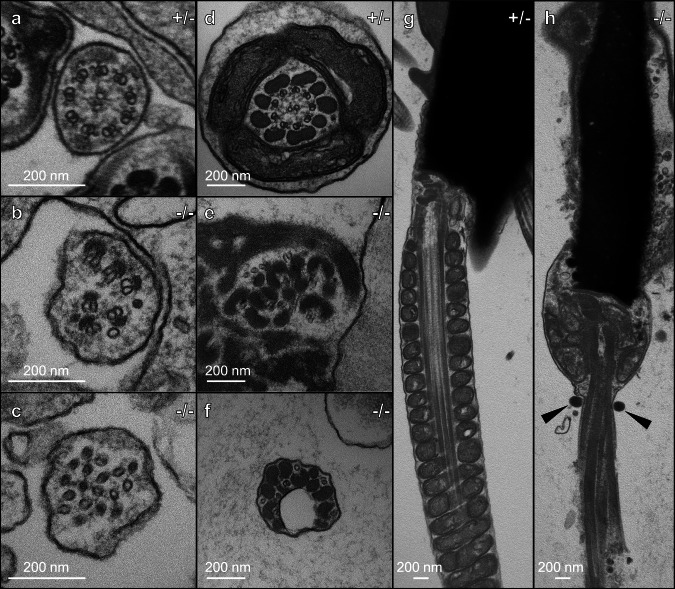




References
-
- Miyata, H., Shimada, K., Kaneda, Y. & Ikawa, M. Development of functional spermatozoa in mammalian spermiogenesis. Development151, dev202838 (2024). - PubMed
-
- Zhou, L. et al. Structures of sperm flagellar doublet microtubules expand the genetic spectrum of male infertility. Cell186, 2897–2910.e19 (2023). - PubMed
MeSH terms
Grants and funding
- P01 HD087157/HD/NICHD NIH HHS/United States
- JP23jf0126001/Japan Agency for Medical Research and Development (AMED)
- JPMJFR211F/MEXT | Japan Science and Technology Agency (JST)
- NA/Takeda Science Foundation
- JP23K18328/MEXT | Japan Society for the Promotion of Science (JSPS)
- INV-001902/GATES/Gates Foundation/United States
- R01HD088412/U.S. Department of Health & Human Services | NIH | Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD)
- JP21H04753/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP23K20043/MEXT | Japan Society for the Promotion of Science (JSPS)
- P01HD087157/U.S. Department of Health & Human Services | NIH | Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD)
- JP21H05033/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP25K02773/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP19H05750/MEXT | Japan Society for the Promotion of Science (JSPS)
- R01 GM129325/GM/NIGMS NIH HHS/United States
- R01 HD088412/HD/NICHD NIH HHS/United States
- JP22H03214/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP23K05831/MEXT | Japan Society for the Promotion of Science (JSPS)
- INV-001902/GATES/Gates Foundation/United States
LinkOut - more resources
Full Text Sources
Research Materials
