

Structural basis for LZTR1 recognition of RAS GTPases for degradation
- PMID: 40934300
- PMCID: PMC12516822
- DOI: 10.1126/science.adv7088
Structural basis for LZTR1 recognition of RAS GTPases for degradation
Abstract
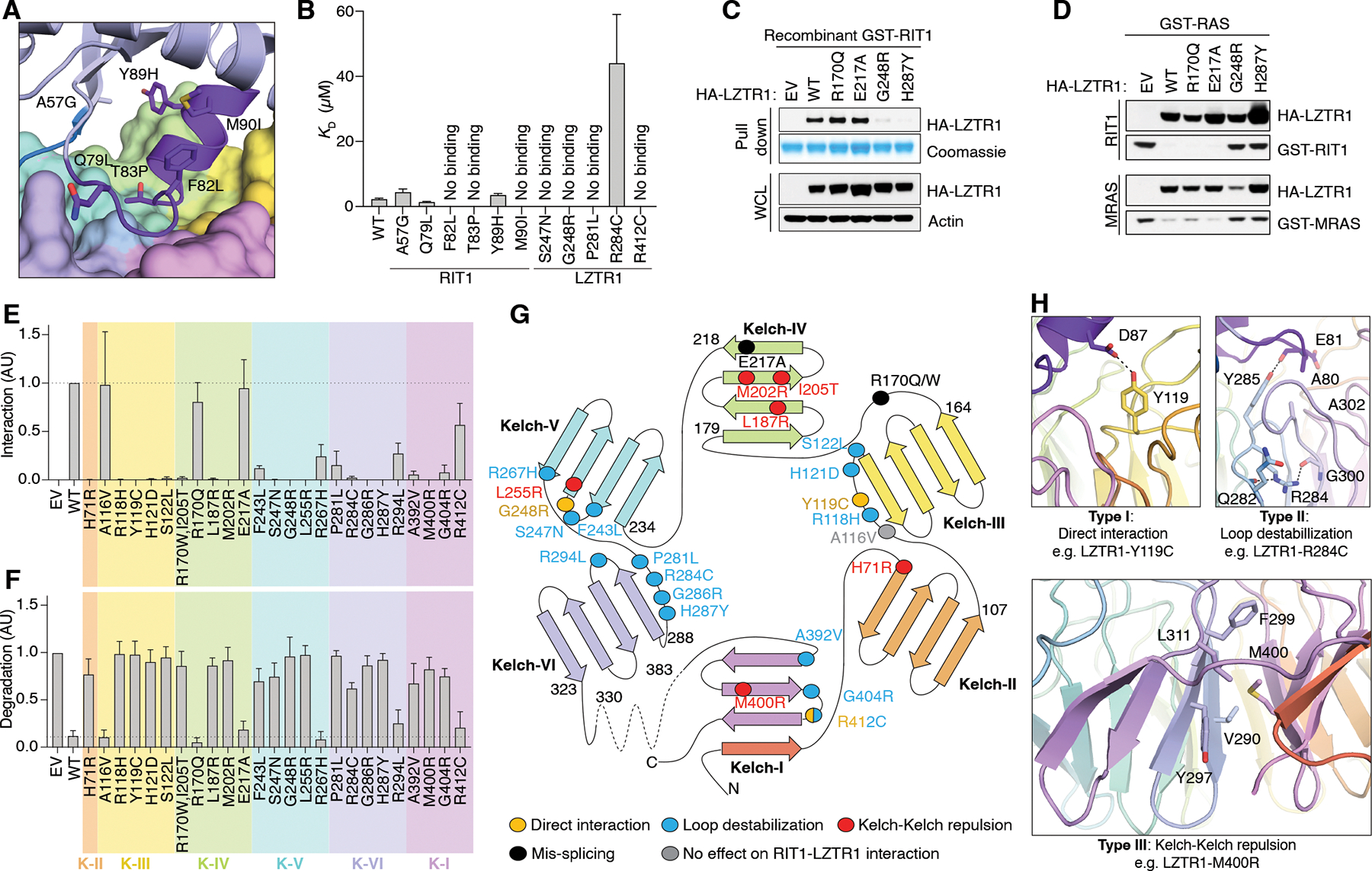
The RAS family of small guanosine triphosphatases (GTPases) are tightly regulated signaling molecules that are further modulated by ubiquitination and proteolysis. Leucine Zipper-like Transcription Regulator 1 (LZTR1), a substrate adapter of the Cullin-3 RING E3 ubiquitin ligase, binds specific RAS GTPases and promotes their ubiquitination and proteasomal degradation. We present structures of LZTR1 Kelch domains bound to RIT1, MRAS, and KRAS, revealing interfaces that govern RAS isoform selectivity and nucleotide specificity. Biochemical and structural analyses of disease-associated Kelch domain mutations revealed three types of alterations: impaired substrate interaction, loop destabilization, and blade-blade repulsion. In cellular and mouse models, mutations disrupting substrate binding phenocopied LZTR1 loss, underscoring its substrate specificity. These findings define RAS recognition mechanisms by LZTR1 and suggest a molecular glue strategy to degrade oncogenic KRAS.
Conflict of interest statement
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
