

Cardiac lymphatics retain LYVE-1-dependent macrophages during neonatal mouse heart regeneration
- PMID: 40962932
- PMCID: PMC12520984
- DOI: 10.1038/s44161-025-00711-4
Cardiac lymphatics retain LYVE-1-dependent macrophages during neonatal mouse heart regeneration
Abstract
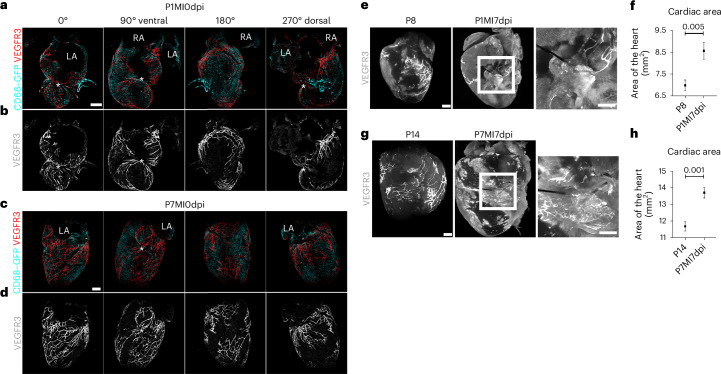
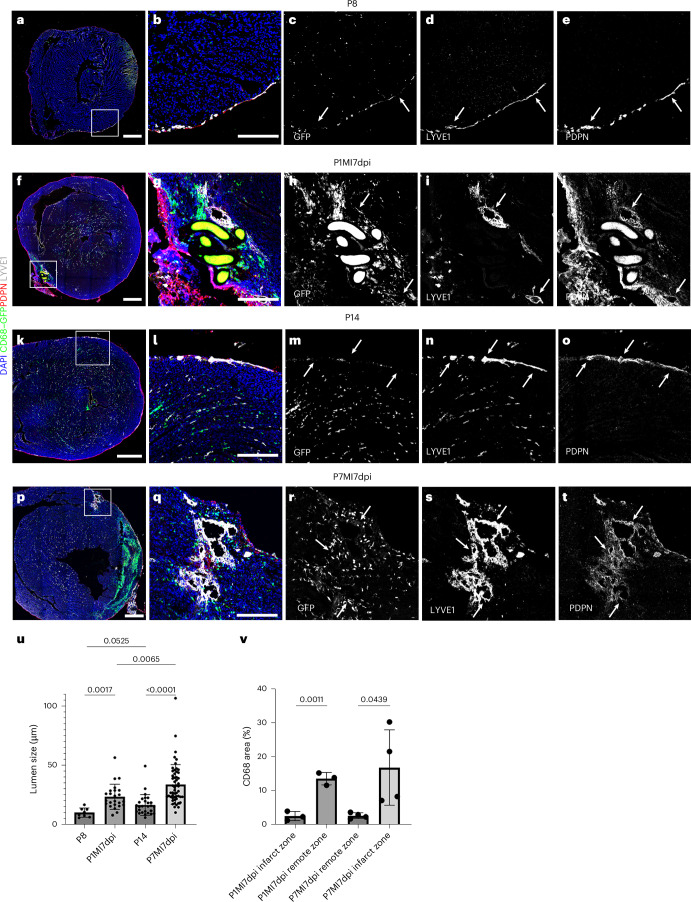
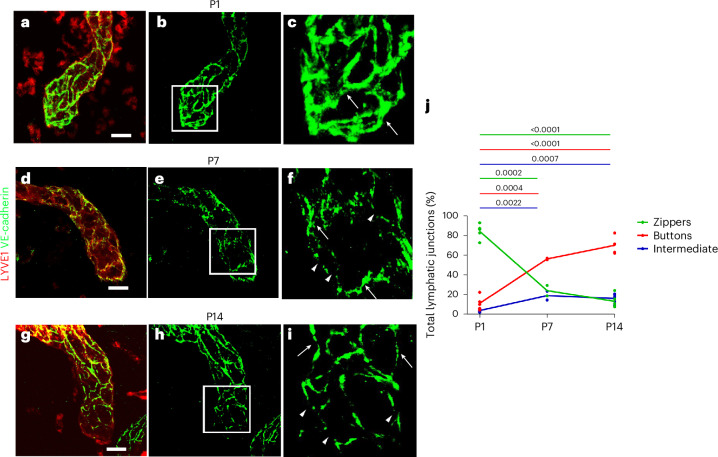
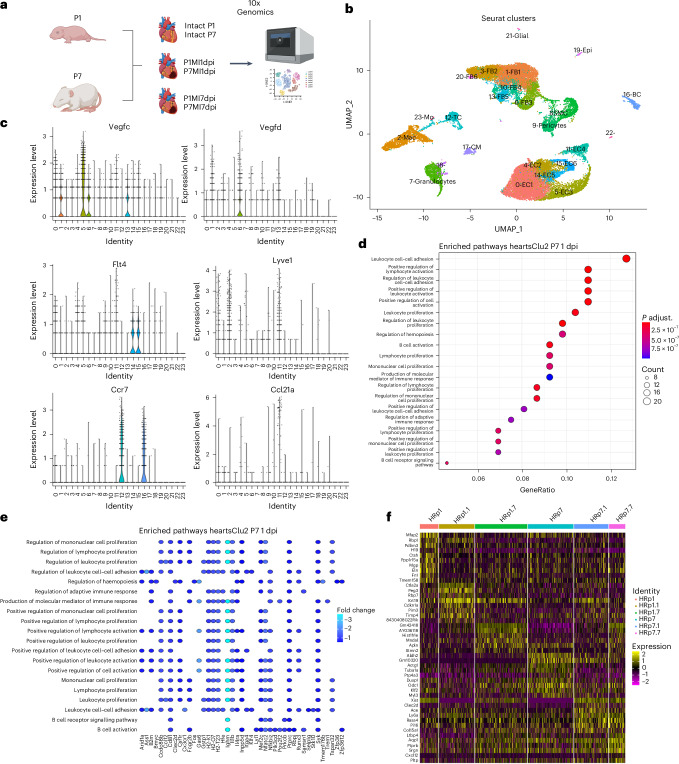
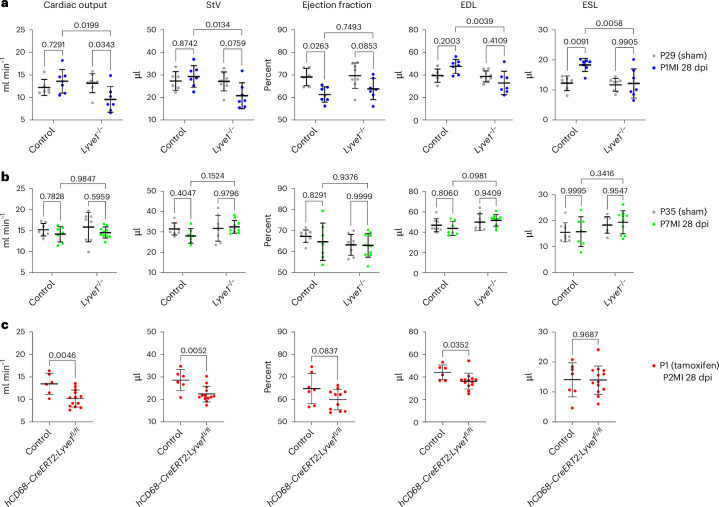
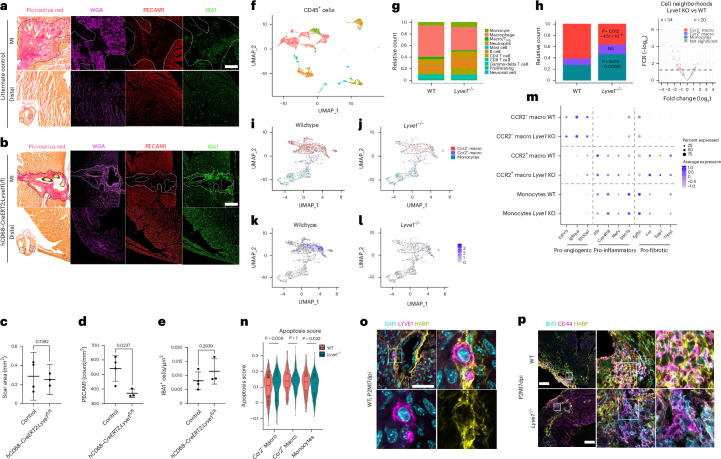
In adult mice, myocardial infarction (MI) activates the cardiac lymphatics, which undergo sprouting angiogenesis (lymphangiogenesis), drain interstitial fluid and traffic macrophages to mediastinal lymph nodes (MLNs). This prevents edema and reduces inflammatory/fibrotic immune cell content to improve cardiac function. Here we investigated the role of cardiac lymphatics and macrophage clearance across the neonatal mouse regenerative window. The response to injury revealed limited lymphangiogenesis and clearance of macrophages from postnatal day 1 compared to postnatal day 7 infarcted hearts. This coincides with the maturation of lymphatic endothelial cell junctions from impermeable to permeable and with altered signaling between lymphatic endothelial cells and macrophages. Mice lacking the lymphatic endothelial receptor-1 (LYVE-1), where macrophage lymphatic trafficking is impaired in adults, experienced worse long-term outcomes after MI induced at postnatal day 1, suggesting an alternative role for LYVE-1 in macrophages. Macrophage-specific deletion of Lyve1 during neonatal heart injury impaired heart regeneration. This study demonstrates that immature cardiac lymphatics are impermeable to clearance in early neonates, ensuring retention of pro-regenerative LYVE-1-dependent macrophages.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures












References
-
- Henri, O. et al. Selective stimulation of cardiac lymphangiogenesis reduces myocardial edema and fibrosis leading to improved cardiac function following myocardial infarction. Circulation133, 1484–1497 (2016). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous