

Continent-wide differentiation of fitness traits and patterns of climate adaptation among European populations of Drosophila melanogaster
- PMID: 40980703
- PMCID: PMC12448211
- DOI: 10.1093/evlett/qraf014
Continent-wide differentiation of fitness traits and patterns of climate adaptation among European populations of Drosophila melanogaster
Abstract
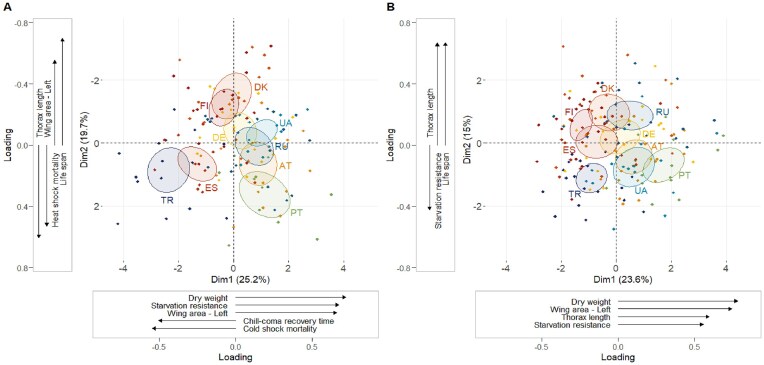
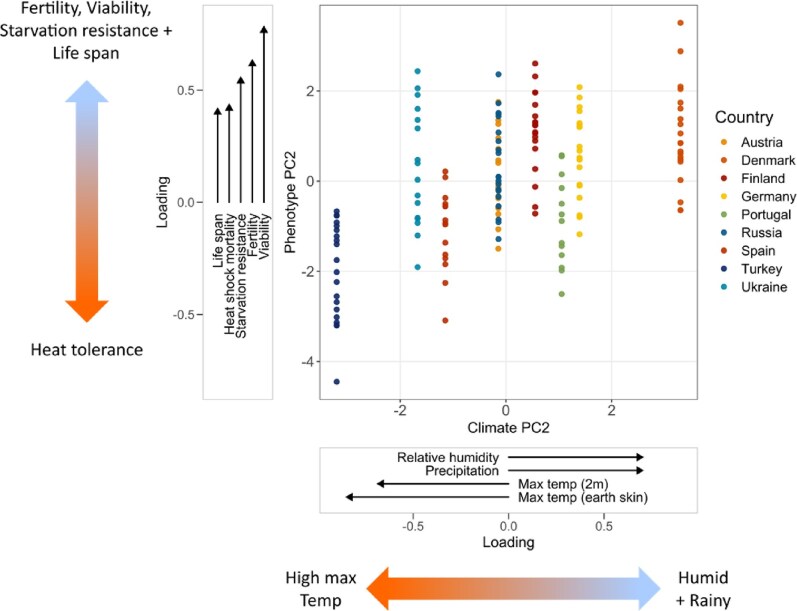
A particularly well-studied evolutionary model is the vinegar fly Drosophila melanogaster, a cosmopolitan insect of ancestral southern-central African origin. Recent work suggests that it expanded out of Africa ∼9,000 years ago, and spread from the Middle East into Europe ∼1,800 years ago. During its global expansion, this human commensal adapted to novel climate zones and habitats. Despite much work on phenotypic differentiation and adaptation on several continents (especially North America and Australia), typically in the context of latitudinal clines, little is known about phenotypic divergence among European populations. Here, we sought to provide a continent-wide study of phenotypic differentiation among European populations of D. melanogaster. In a consortium-wide phenomics effort, we assayed 16 fitness-related traits on a panel of 173 isofemale lines from 9 European populations, with the majority of traits measured by several groups using semi-standardized protocols. For most fitness-related traits, we found significant differentiation among populations on a continental scale. Despite inevitable differences in assay conditions among labs, the reproducibility and hence robustness of our measurements were overall remarkably good. Several fitness components (e.g., viability, development time) exhibited significant latitudinal or longitudinal clines, and populations differed markedly in multivariate trait structure. Notably, populations experiencing higher humidity/rainfall and lower maximum temperature showed higher viability, fertility, starvation resistance, and lifespan at the expense of lower heat-shock survival, suggesting a pattern of local adaptation. Our results indicate that derived populations of this tropical fly have been shaped by pervasive spatially varying multivariate selection and adaptation to different climates on the European continent.
Keywords: D. melanogaster; Europe; adaptation; fitness traits; phenotypic variation; population differentiation.
© The Author(s) 2025. Published by Oxford University Press on behalf of The Society for the Study of Evolution (SSE) and European Society for Evolutionary Biology (ESEB).
Conflict of interest statement
The authors declare no conflict of interest. Note: After February 24, 2022, no collaborative actions or exchanges have taken place between Ukrainian and Russian scientists within our project.
Figures


References
-
- Ackermann M., Bijlsma R., James A. C., Partridge L., Zwaan B. J., Stearns S. C. (2001). Effects of assay conditions in life history experiments with Drosophila melanogaster . Journal of Evolutionary Biology, 14:(2), 199–209. 10.1046/j.1420-9101.2001.00281.x - DOI
LinkOut - more resources
Full Text Sources
