

Development of clinical immunity to Plasmodium vivax following repeat controlled human malaria infection
- PMID: 40998768
- PMCID: PMC12462441
- DOI: 10.1038/s41467-025-63104-y
Development of clinical immunity to Plasmodium vivax following repeat controlled human malaria infection
Abstract
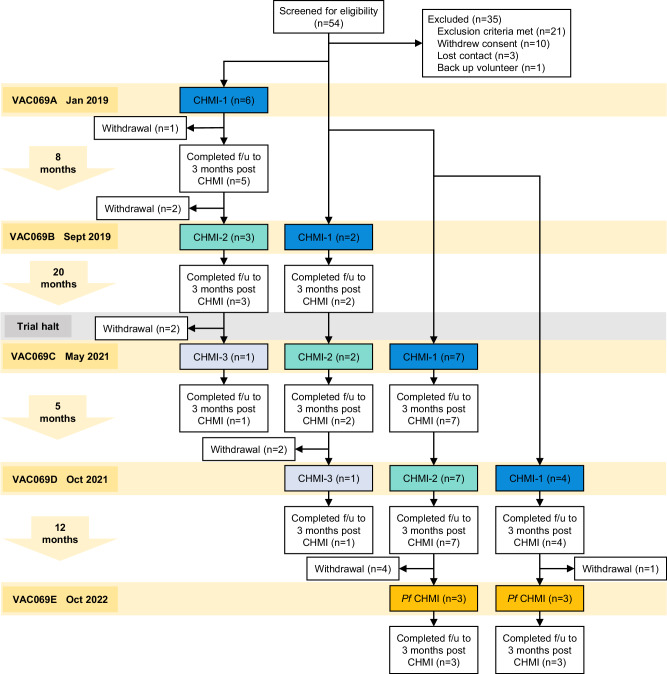
Clinical immunity to malaria can lead to asymptomatic infection, but the underlying mechanisms remain unclear. To examine the development of clinical immunity, we conducted a multi-cohort, repeat controlled human malaria infection (CHMI) study with Plasmodium vivax, and a heterologous rechallenge with P. falciparum (ClinicalTrials.gov NCT03797989). Malaria-naïve adults underwent CHMI up to three times, by administration of red blood cells infected with P. vivax PvW1 clone or P. falciparum 3D7 clone. Nineteen participants underwent primary CHMI with P. vivax, 12 returned for secondary homologous CHMI and 2 for tertiary homologous CHMI. Six participants who had completed P. vivax CHMI then underwent heterologous rechallenge with P. falciparum. We find that clinical immunity to P. vivax develops rapidly after a single CHMI, protecting participants against fever and laboratory abnormalities. This is underpinned by the attenuation of inflammatory cytokines and chemokines, as well as reduced coagulation and endothelium activation. In contrast, there is no evidence of anti-parasite immunity, suggesting that mechanisms of clinical immunity can operate independently of pathogen load to reduce the damage caused by malaria infection. In addition, we show that clinical immunity to P. vivax is parasite species-specific and provides no protection against CHMI with P. falciparum.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures




References
-
- Word Health Organisation. World Malaria Report 2024: Addressing Inequity in the Global Malaria Response (2024).
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
