

PA2G4 Functions as a Cofactor for MYC Family Oncoproteins in MYC-Driven Malignancies
- PMID: 41002386
- PMCID: PMC12468391
- DOI: 10.3390/cells14181422
PA2G4 Functions as a Cofactor for MYC Family Oncoproteins in MYC-Driven Malignancies
Abstract
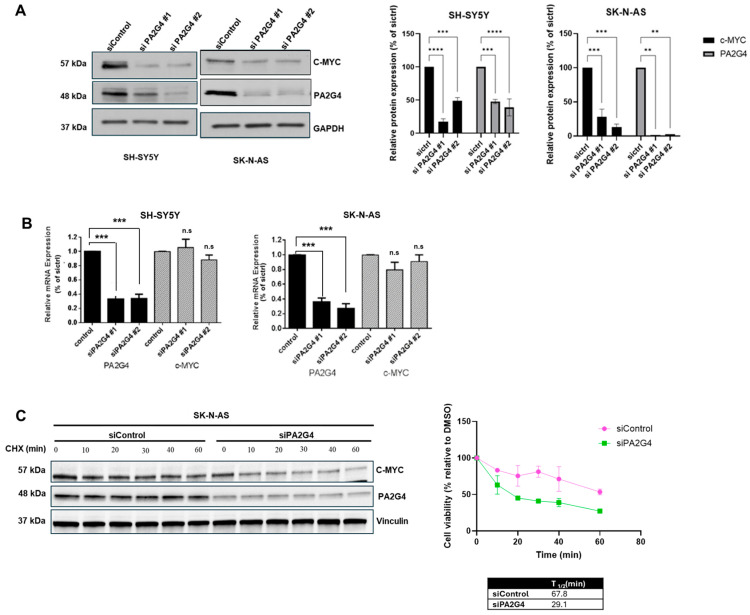
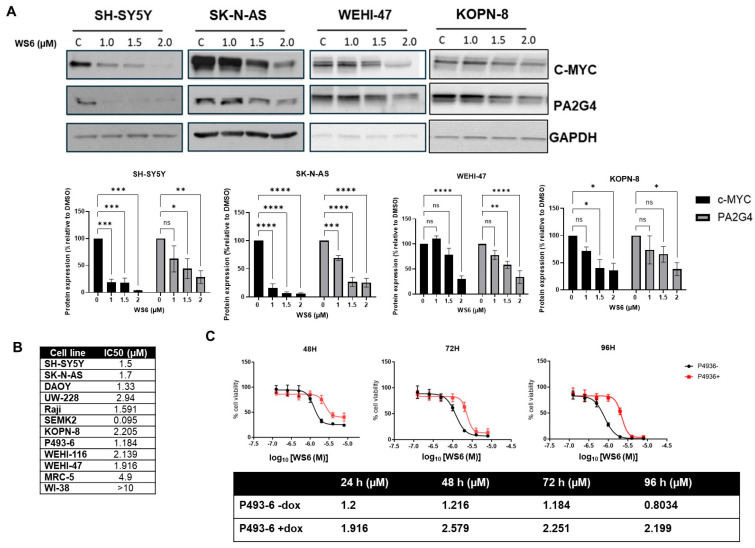
MYCN and c-MYC are critical driver oncogenes in several childhood cancers, including neuroblastoma. Currently, the clinical development of MYC inhibitors has been hindered by the intrinsically disordered structure of MYC proteins, which lack well-defined ligand-binding pockets. Proliferation-associated protein 2G4 (PA2G4) directly binds to and stabilizes MYCN protein, leading to markedly increased MYCN levels in neuroblastoma cells. Here, we demonstrate that PA2G4 is essential for MYCN-driven tumor growth in neuroblastoma in vivo. Moreover, PA2G4 elevates c-MYC protein levels in neuroblastoma cells by inhibiting its ubiquitin-mediated degradation. In turn, c-MYC upregulates the transcription and protein expression of PA2G4, creating an oncogenic feed-forward expression loop. A small molecule PA2G4 inhibitor, WS6, directly disrupts the PA2G4-c-MYC protein-protein interaction, resulting in decreased levels of both PA2G4 and c-MYC. WS6 exhibited selective cytotoxicity in c-MYC-overexpressing cell lines. Together, these findings identify PA2G4 as a shared cofactor for both the c-MYC and MYCN oncoproteins and highlight its interaction with MYC family oncoproteins as a promising therapeutic vulnerability in MYC-driven cancers.
Keywords: Burkitt’s lymphoma; MYC; MYCN; PA2G4; WS6; inhibitors; neuroblastoma.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures



References
-
- Chou Y.T., Lin H.H., Lien Y.C., Wang Y.H., Hong C.F., Kao Y.R., Lin S.C., Chang Y.C., Lin S.Y., Chen S.J., et al. EGFR promotes lung tumorigenesis by activating miR-7 through a Ras/ERK/Myc pathway that targets the Ets2 transcriptional repressor ERF. Cancer Res. 2010;70:8822–8831. doi: 10.1158/0008-5472.CAN-10-0638. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases