

A basic framework to explain splice-site choice in eukaryotes
- PMID: 41022769
- PMCID: PMC12480690
- DOI: 10.1038/s41467-025-63622-9
A basic framework to explain splice-site choice in eukaryotes
Abstract
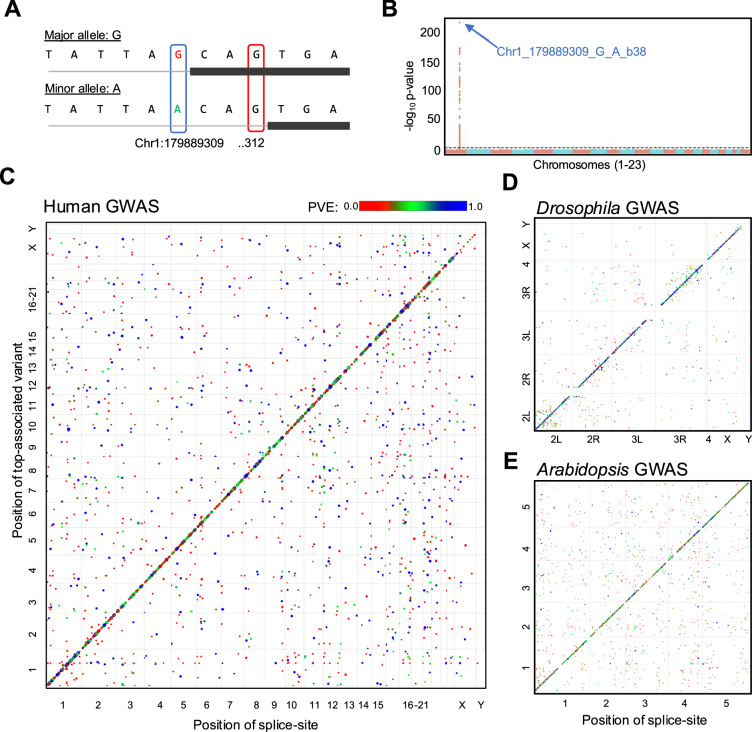
Changes in splicing can mediate phenotypic variation, ranging from flowering time differences in plants to genetic diseases in humans. Splicing changes occur due to differences in splice-site strength, often influenced by genetic variation and the environment. How genetic variation influences splice-site strength remains poorly understood, largely because splice-site usage across transcriptomes has not been empirically quantified. Here, we quantify the use of individual splice-sites in Arabidopsis, Drosophila and humans and treat these measurements as molecular phenotypes to map variation in splice-site usage through GWAS. We carry out more than 130,000 GWAS with splice-site usage phenotypes, cataloguing genetic variation associated with changes in the usage of individual splice-sites across transcriptomes. We find that most of the common, genetically controlled variation in splicing is cis and there are no major trans hotspots in the three species analyzed. We group splice-sites based on GT[N]4 or [N]4AG sequence, quantify their average use, develop a ranking and show that these hexamer rankings provide a simple and comparable feature across species to explain most of the splice-site choice. Transcriptome analyses in several species suggest that hexamer rankings offer a rule that helps explain splice-site choices, forming a basis for a shared splicing logic in eukaryotes.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



References
-
- Rogalska, M. E., Vivori, C. & Valcarcel, J. Regulation of pre-mRNA splicing: roles in physiology and disease, and therapeutic prospects. Nat. Rev. Genet.24, 251–269 (2023). - PubMed
-
- Marasco, L. E. & Kornblihtt, A. R. The physiology of alternative splicing. Nat. Rev. Mol. Cell Biol.24, 242–254 (2023). - PubMed
-
- Nasif, S., Contu, L. & Muhlemann, O. Beyond quality control: The role of nonsense-mediated mRNA decay (NMD) in regulating gene expression. Semin. Cell Dev. Biol.75, 78–87 (2018). - PubMed
-
- Black, D. L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem.72, 291–336 (2003). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
