

Cholecystokinin ameliorates cognitive impairment via inhibiting microglia phagocytosis of excitatory synapses in sepsis-associated encephalopathy mice
- PMID: 41024113
- PMCID: PMC12482099
- DOI: 10.1186/s12974-025-03554-9
Cholecystokinin ameliorates cognitive impairment via inhibiting microglia phagocytosis of excitatory synapses in sepsis-associated encephalopathy mice
Abstract
Background: Sepsis-associated encephalopathy (SAE) is characterised by cognitive impairment and is a common complication in patients with sepsis. Microglia are involved in various cognitive impairment-related diseases through phagocytic synapses. Cholecystokinin (CCK), an abundant neuropeptide in the brain, is closely related to cognitive function. However, the role of CCK in SAE and the relationship between CCK and microglial phagocytosis of synapses are unknown.
Methods: Lipopolysaccharide (LPS) was used to construct SAE models in 3-month-old male mice and BV2 microglial cells. To investigate the effects of CCK on cognitive impairment in SAE model mice, we used exogenous CCK injection into the dorsal hippocampal CA1 region or the chemogenetic activation of CCK-positive neurons to promote endogenous CCK release. Morris water maze and fear conditioning test were used to assess cognitive function in mice. RNA sequencing was performed to explore the potential signalling pathways involved in CCK-induced neuroprotection. Western blot and immunofluorescence were used to assess the effects of CCK on microglial phagocytosis of synapses, neurotoxic astrocytes, and excitatory synapses. Whole-cell recording was used to determine excitatory synaptic transmission.
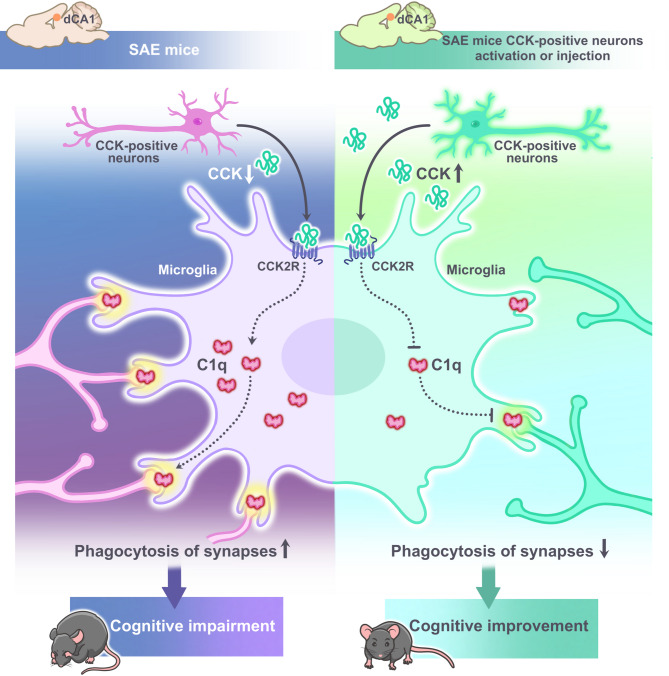
Results: LPS successfully established in vivo and in vitro models of SAE. Both exogenous CCK injection and activation of CCK-positive neurons in hippocampal CA1 region attenuated cognitive impairment in SAE mice. Mechanistically, CCK significantly alleviated excitatory synaptic plasticity damage via inhibiting complement 1q (C1q)-mediated microglial phagocytosis of synapses and neurotoxic astrocyte polarisation. Moreover, in vitro SAE model of BV2 cells demonstrated that CCK exerts neuroprotective effects through microglial CCK2-type receptor.
Conclusions: CCK may alleviate cognitive impairment by inhibiting microglia C1q-mediated phagocytosis of excitatory synapses, suggesting that both CCK drugs and specific activation of CCK-positive neurons are potential treatments for SAE.
Keywords: Cholecystokinin; Complement 1q; Excitatory synapse; Microglia; Sepsis-associated encephalopathy.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: All animal care and experimental procedures were approved by the animal ethics committee of Peking University Health Science Center (ethics number: LA2021534). Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures






References
-
- Gofton TE, Young GB. Sepsis-associated encephalopathy. Nat Rev Neurol. 2012;8(10):557–66. - PubMed
-
- Golzari SE, Mahmoodpoor A. Sepsis-associated encephalopathy versus sepsis-induced encephalopathy. Lancet Neurol. 2014;13(10):967–8. - PubMed
-
- Catarina AV, Branchini G, Bettoni L, De Oliveira JR, Nunes FB. Sepsis-associated encephalopathy: from pathophysiology to progress in experimental studies. Mol Neurobiol. 2021;58(6):2770–9. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
