

Plant-derived vesicle-like nanoparticles for immunomodulation: Mechanisms and applications
- PMID: 41035428
- PMCID: PMC12481952
- DOI: 10.1016/j.bioactmat.2025.09.024
Plant-derived vesicle-like nanoparticles for immunomodulation: Mechanisms and applications
Abstract
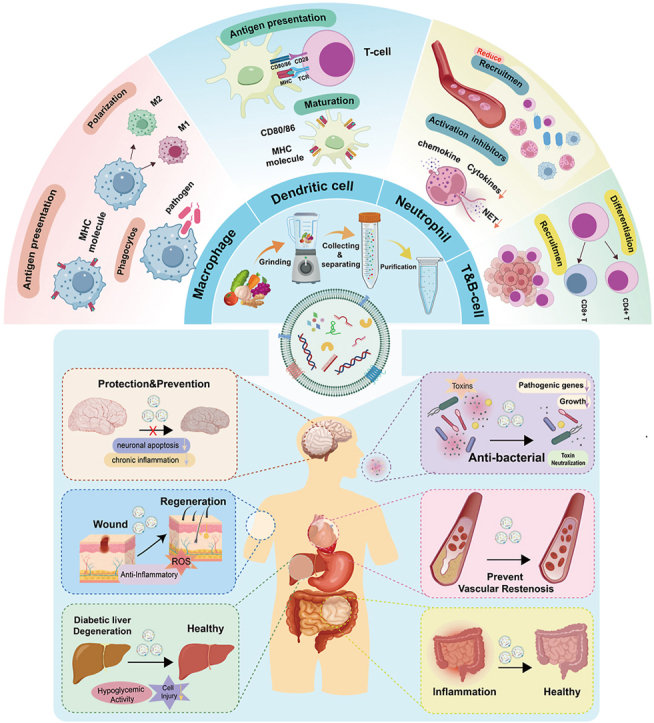
Immune dysregulation can result in sustained activation of the immune system, leading to systemic chronic inflammation. This condition significantly disrupts immune homeostasis and is intimately associated with the onset of numerous chronic systemic diseases. Currently, the treatment of diseases related to immune dysregulation confronts several challenges, most notably the substantial side effects and inconsistent efficacy of long-term immunosuppressive drug use. Consequently, developing immunomodulatory strategies that balance efficacy and safety has emerged as a prominent research focus. Plant-derived vesicle-like nanoparticles (PVLNs), natural nanomaterials secreted by plant cells, exhibit significant potential in immunomodulation owing to their excellent biocompatibility, minimal immunogenicity, and cross-species communication capabilities. This paper reviews the biogenesis, composition, and properties of PVLNs, emphasizing the mechanisms of innate and adaptive immunomodulation they mediate and their applications in diseases characterized by immune disorders. It also analyzes the challenges related to target delivery, stability optimization, drug loading, and storage encountered in their engineering. In the future, as the mechanisms of PVLNs are more deeply understood and nanotechnology continues to advance, their potential in precision immunotherapy and clinical translation is anticipated to be further augmented.
Keywords: Engineered modifications; Immunomodulation; Natural nanomaterials; Plant vesicles.
© 2025 The Authors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures












References
-
- Furman D., Campisi J., Verdin E., Carrera-Bastos P., Targ S., Franceschi C., Ferrucci L., Gilroy D.W., Fasano A., Miller G.W., Miller A.H., Mantovani A., Weyand C.M., Barzilai N., Goronzy J.J., Rando T.A., Effros R.B., Lucia A., Kleinstreuer N., Slavich G.M. Chronic inflammation in the etiology of disease across the life span. Nat Med. 2019;25:1822–1832. doi: 10.1038/s41591-019-0675-0. - DOI - PMC - PubMed