

Dynamic neural compensation for distorted orientation perception in chronic astigmatism
- PMID: 41079638
- PMCID: PMC12510184
- DOI: 10.1016/j.isci.2025.113518
Dynamic neural compensation for distorted orientation perception in chronic astigmatism
Abstract
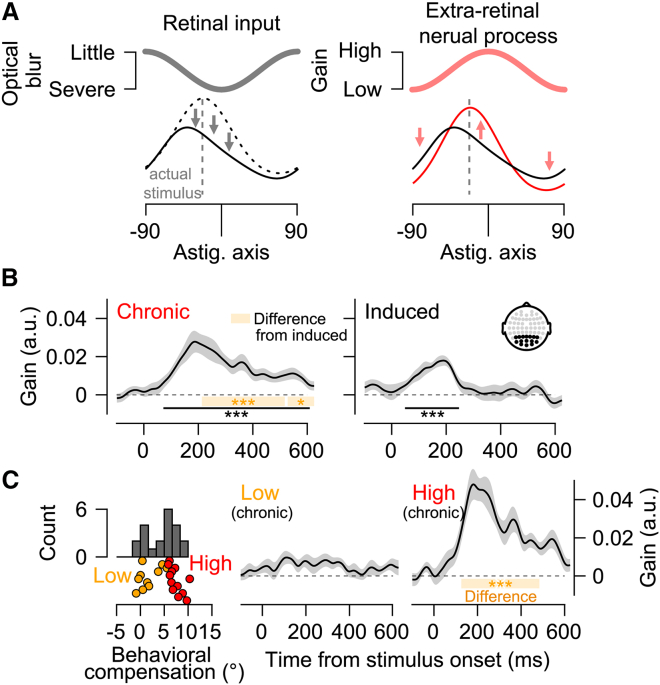
Astigmatism induces orientation-specific blur, distorting retinal inputs, and biasing perception. Yet individuals with chronic astigmatism often perceive orientation accurately, though the underlying neural mechanisms remain unclear. We investigated how the brain compensates for this distortion by recording electroencephalogram (EEG) responses from 42 participants (15 females) under astigmatic viewing conditions. Using multivariate EEG decoding and a computational model, we reconstructed population orientation tuning curves and identified neural modulation not explained by retinal inputs alone. Chronic astigmatism enhanced neural sensitivity to blurred orientations and suppressed responses to intact ones, forming a push-pull modulation pattern correlated with perceptual accuracy. This modulation dynamically propagated from posterior to anterior brain regions after stimulus onset and predicted the perceptual compensation. In contrast, short-term exposure produced weak, transient modulation, and an opposite anterior-to-posterior propagation pattern without perceptual relevance. These findings show that long-term visual distortion reshapes neural tuning and inter-regional dynamics, enabling perceptual optimization through cortical plasticity.
Keywords: Neuroscience; Sensory neuroscience.
© 2025 The Authors.
Conflict of interest statement
Authors declare that they have no competing interests.
Figures






References
LinkOut - more resources
Full Text Sources
