

Targeting PURPL RNA enabled rejuvenation of senescence cells via epigenetic reprogramming
- PMID: 41107828
- PMCID: PMC12535130
- DOI: 10.1186/s12967-025-07208-5
Targeting PURPL RNA enabled rejuvenation of senescence cells via epigenetic reprogramming
Abstract
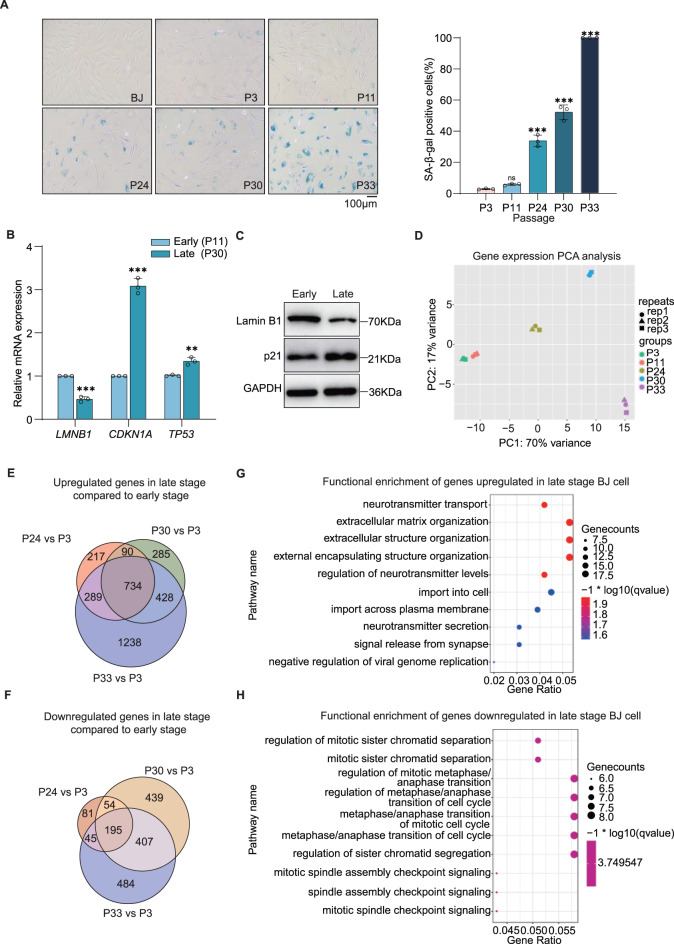
Cellular senescence is a fundamental driver of ageing and age-related diseases, characterized by irreversible growth arrest and profound epigenetic alterations. While long non-coding RNAs (lncRNAs) have emerged as key regulators of senescence, their potential for senescent cell rejuvenation remains unexplored. Here, we identify the ageing-associated lncRNA PURPL as an epigenetic regulator that controls cellular rejuvenation through H3K9me3-mediated transcriptional silencing. CRISPRi-mediated PURPL depletion produces striking rejuvenation effects, resulting in restored youthful cell morphology, as well as suppression of senescence markers such as p21 and SA-β-gal. Conversely, PURPL overexpression accelerates cellular senescence, recapitulating the transcriptional and phenotypic hallmarks of ageing. Mechanistically, nuclear-localized PURPL regulates H3K9me3 deposition at 411 genomic loci including SERPINE1 (PAI-1) and EGR1, which are key senescence drivers. PURPL-mediated H3K9me3 loss at these loci derepresses their transcription, establishing a pro-senescence gene expression program. These findings reveal that PURPL is an epigenetic modulator of senescence and highlight its potential as a therapeutic target for age-related pathologies.
Keywords: Cellular senescence; H3K9me3; Histone modification; Long non-coding RNA; PURPL.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures




References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
