

Trained Immunity in Health and Disease
- PMID: 41211186
- PMCID: PMC12592693
- DOI: 10.1002/mco2.70461
Trained Immunity in Health and Disease
Abstract
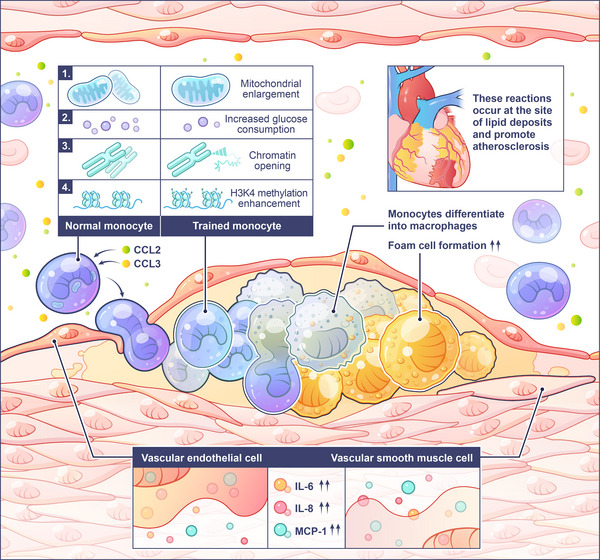
Trained immunity as a critical regulator of host defense and disease pathogenesis bridges the gap between innate and adaptive immunity. For decades, the classic dichotomy of innate immunity and adaptive immunity has shaped our knowledge of immune function. Innate immunity has traditionally been regarded as a rapid, nonspecific first line of defense without memory capacity, while adaptive immunity is characterized by slower, antigen-specific responses and long-term immune memory. However, emerging evidence that innate immunity exhibits memory-like properties challenges the paradigm. Basically, innate immune cells with nonspecific memory retain functional imprints of prior encounters with diverse stimuli. Here, we comprehensively explore the intricate molecular and cellular mechanisms that underpin trained immunity, encompassing epigenetic inheritance, metabolic reprogramming, and transcriptional rewiring. Its dual roles are highlighted in health and disease. On one hand, it bolsters host defense against a broad spectrum of pathogens from bacteria to viruses, and enhances vaccine efficacy through heterologous protection. On the other hand, its dysregulation contributes to infection, inflammation, and cancer progression. As for the promising opportunities on therapeutic intervention, the challenges in precisely modulating trained immunity are tackled to offer a holistic perspective on the dynamically evolving field.
Keywords: epigenetic inheritance; immune cells; immunotherapy; metabolic reprogramming; trained immunity.
© 2025 The Author(s). MedComm published by Sichuan International Medical Exchange & Promotion Association (SCIMEA) and John Wiley & Sons Australia, Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures



References
-
- Katzmarski N., Domínguez‐Andrés J., Cirovic B., et al., “Transmission of Trained Immunity and Heterologous Resistance to Infections Across Generations,” Nature Immunology 22, no. 11 (2021): 1382–1390. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous