

This is a preprint.
Mutant MAPT Induces rDNA Transcriptional Hyperactivation and Nucleolar Stress in Cellular Models
- PMID: 41255983
- PMCID: PMC12622150
- DOI: 10.21203/rs.3.rs-7713987/v1
Mutant MAPT Induces rDNA Transcriptional Hyperactivation and Nucleolar Stress in Cellular Models
Abstract
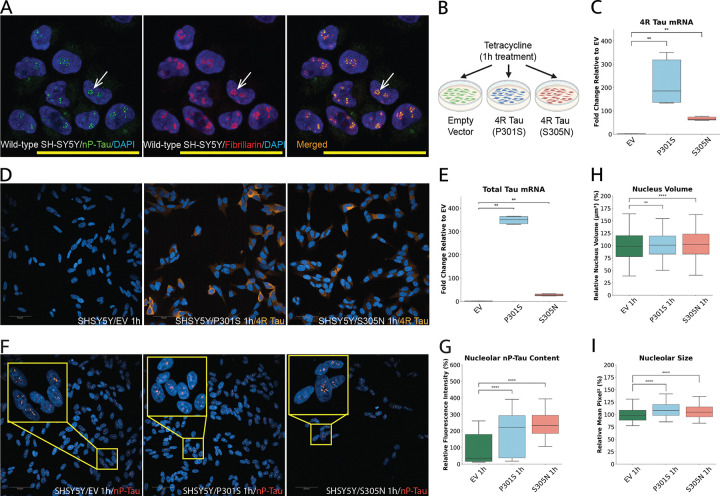
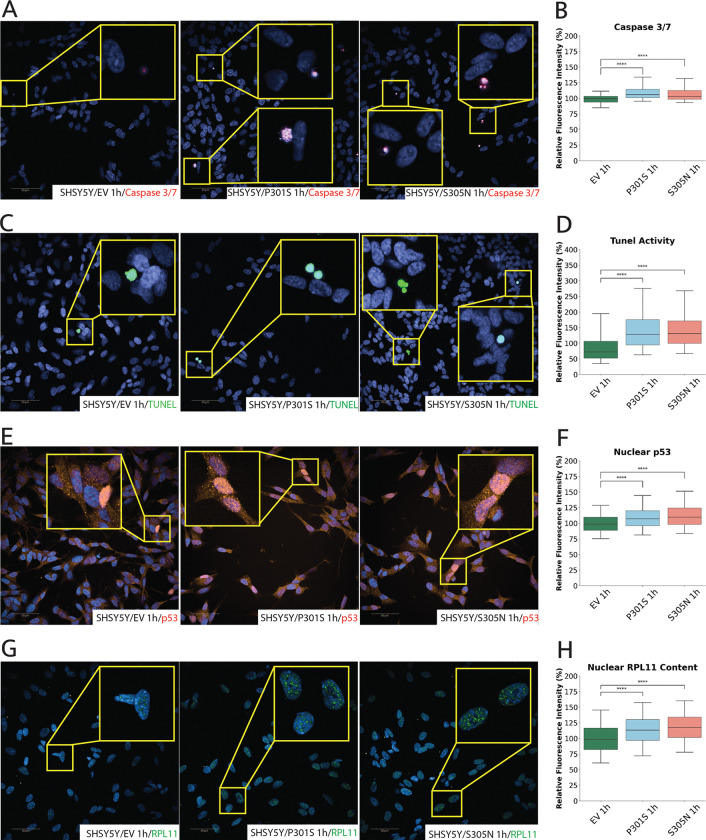
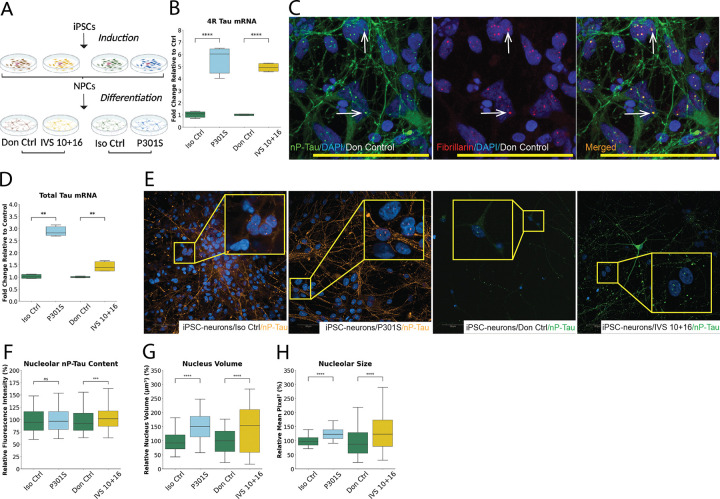
Tau is traditionally known for its role in microtubule stabilization, with its pathological aggregation central to tauopathies such as Alzheimer's disease (AD) and frontotemporal dementia (FTD). Recent evidence suggests that tau also plays important nuclear and nucleolar roles, yet the implications of tau pathology on nucleolar function remain poorly understood. Here, we show that tau localises to the nucleolus in both differentiated SH-SY5Y cells and iPSC-derived neurons, and accumulates upon expression of disease-associated MAPT mutations (P301S, S305N, and IVS 10 + 16). Using high-content imaging, we demonstrate that mutant tau expression leads to structural expansion of the nucleus and nucleolus, with upregulation of key markers from all three nucleolar sub-compartments, indicating increased in nucleolar activity. qPCR and nucleolar RNA-selective dye staining confirmed increased rDNA transcription and rRNA processing, suggesting that mutant tau drives elevated nucleolar biosynthetic output. This hyperactivation is accompanied by hallmarks of nucleolar stress and apoptosis, including p53 stabilisation, caspase 3/7 activation, and TUNEL positivity. These findings identify nucleolar dysfunction as a downstream consequence of mutant tau expression and highlight disruption of nucleolar homeostasis as a potential contributor to tau-mediated neurotoxicity in MAPT-linked FTD.
Conflict of interest statement
Declarations Conflict of Interest: All authors declare no competing financial or non-financial interests in relation to the work described in this manuscript.
Figures



References
-
- Brion J. P., Couck A. M., Passareiro E. & Flament-Durand J. Neurofibrillary tangles of Alzheimer’s disease: an immunohistochemical study. J Submicrosc Cytol 17, 89–96 (1985). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
