

This is a preprint.
Coordinated circulating and tissue-based T cell responses precede xenograft rejection
- PMID: 41279278
- PMCID: PMC12637509
- DOI: 10.1101/2025.11.07.687259
Coordinated circulating and tissue-based T cell responses precede xenograft rejection
Abstract
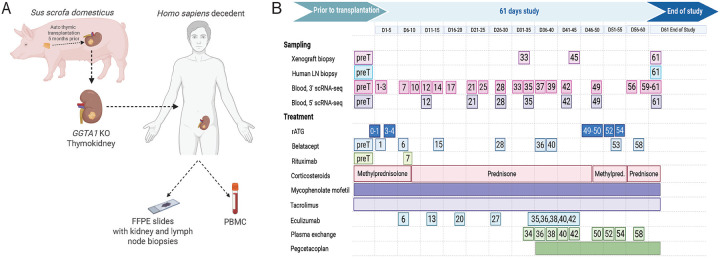
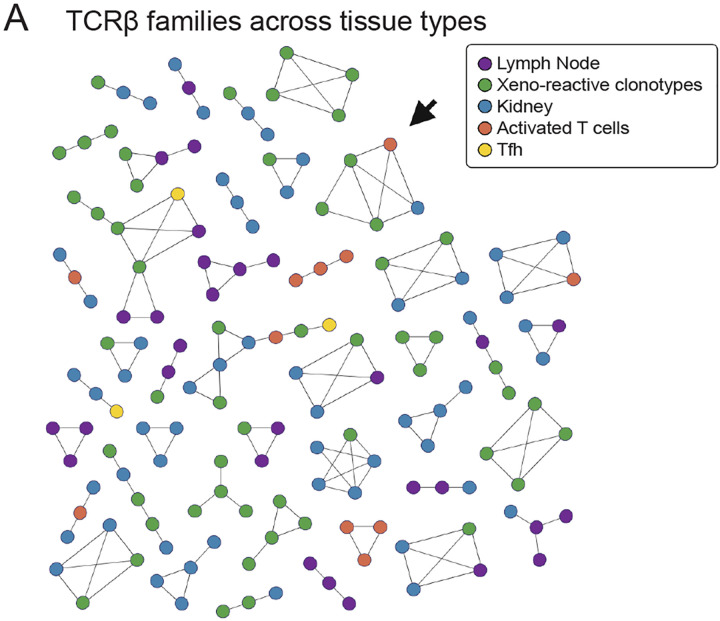
Despite the life-saving successes of solid organ transplantation, the number of individuals needing organ transplant far exceeds the number of organs available for use each year. Porcine xenotransplantation, or the use of pig organs for transplantation in people, holds substantial promise but xenograft rejection in humans is poorly understood. T cell rejection by the host immune system is a major challenge for human allografts and may limit the longevity of porcine xenografts. To study the xenograft rejection, we evaluated T cell responses and repertoire dynamics across tissues following porcine thymokidney transplantation in a decedent model over 61 days after bilateral native kidney nephrectomy. Despite induction with anti-thymocyte globulin and ongoing immune suppression consisting of rituximab, corticosteroids, calcineurin inhibition, and mycophenolate mofetil, human T cell infiltration of the xenograft was observed and was associated with xenograft dysfunction. Longitudinal analysis of T cell clonotypes in biopsies of thymokidney revealed accumulation of clonal human CD4 and CD8 T cell responses. Moreover, circulating activated T cells, including circulating T follicular helper (cTfh), were xeno-reactive and increased in frequency around rejection events. We confirmed clonal dominance of a single CD8 clonotype - identified as donor-reactive in a mixed lymphocyte reaction - in the circulation leading up to the acute cellular rejection event. Following re-treatment with anti-thymocyte globulin and intensification of corticosteroids, the T cell clonotypes were dramatically diminished in frequency in thymokidney and lymph nodes, though not eliminated. Over time of observation, the T cell clonotypes were shared across multiple compartments, including xenograft, circulation and lymph nodes and formed clonal families with known xeno-reactive clonotypes, suggesting a coordinated immune response against a limited pool of antigenic targets. Together, these data demonstrate T cell repertoire dynamics across tissues in the setting of xenograft rejection and highlight opportunities for early surveillance, prediction and potential intervention.
Conflict of interest statement
Competing interests M.S. holds patents #US-5658564-A and #EP-0697876-B1 “Xenograft thymus”.
Figures



References
-
- Health Resources and Services Administration. Organ procurement and transplantation network: national data. 2025.
-
- Pan W, Zhang W, Zheng B, Camellato BR, Stern J, Lin Z, et al. Cellular dynamics in pig-to-human kidney xenotransplantation. Med (N Y). 2024. Aug 9;5(8):1016–1029.e4.
-
- Schmauch E, Piening B, Mohebnasab M, Xia B, Zhu C, Stern J, et al. Integrative multi-omics profiling in human decedents receiving pig heart xenografts. Nat Med. 2024. May 17;30(5):1448–60. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials