

This is a preprint.
Molecular basis for CSB stimulation of the SNM1A DNA repair nuclease
- PMID: 41333433
- PMCID: PMC12668182
- DOI: 10.21203/rs.3.rs-7886236/v1
Molecular basis for CSB stimulation of the SNM1A DNA repair nuclease
Abstract
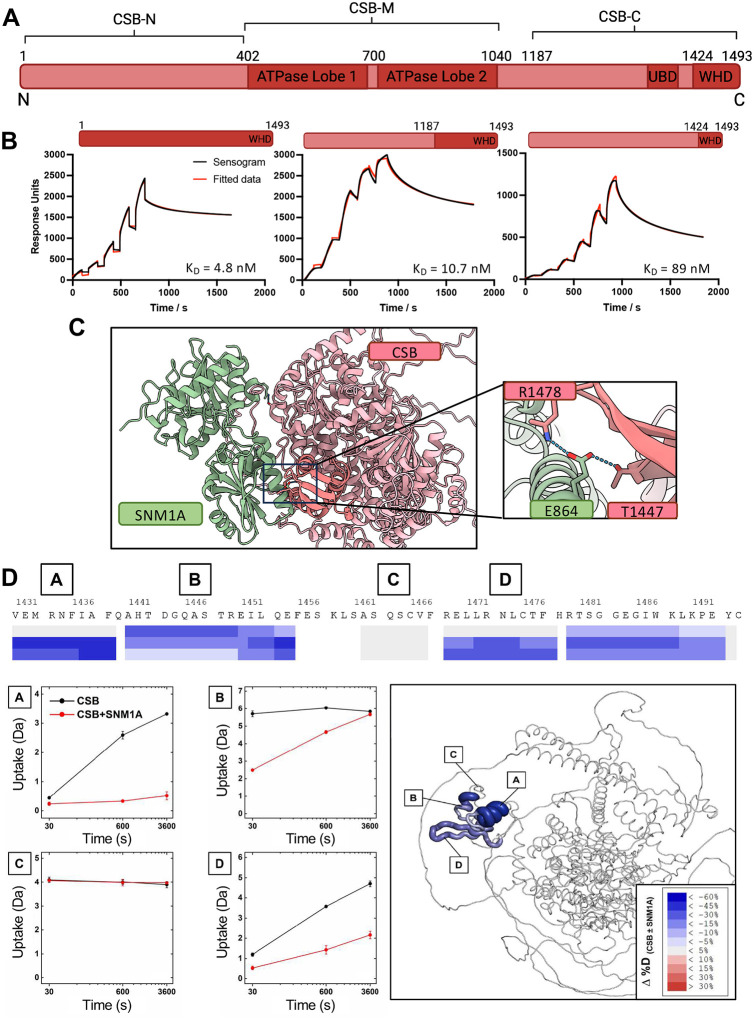
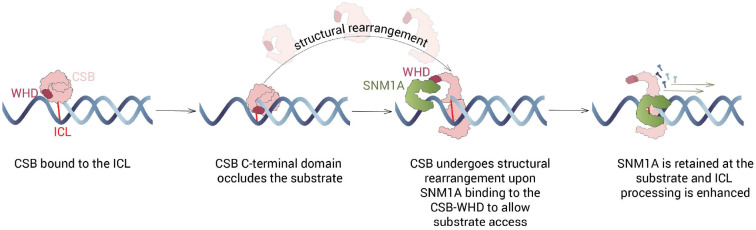
The Cockayne Syndrome B (CSB, ERCC6) protein, interacts with the exonuclease SNM1A during transcription-coupled DNA interstrand (ICL) repair, with CSB facilitating localisation of SNM1A to ICL damage. The functional and mechanistic details of this interaction in DNA repair, however, have not been defined. Here, we demonstrate that CSB enhances SNM1A resection through ICLs and identify a specific interaction between the winged-helix domain of CSB and the nuclease core of SNM1A that is crucial for recruitment and enhancement of nuclease degradation. Biochemical and single-molecule studies on DNA containing sitespecific ICLs reveal that CSB increases the affinity of SNM1A to damaged DNA substrates and also alters the substrate conformation to enhance ICL processing by SNM1A. Notably, CSB was observed preferentially as a dimer when colocalised with SNM1A at ICLs, constrasting with its monomeric nature observed during repair initiation in classical transcription-coupled nucleotide excision repair. The combined results provide molecular insights into the basis of a direct contribution of CSB to a DNA repair reaction.
Figures





References
-
- Clauson C., Schärer O. D. & Niedernhofer L. Advances in understanding the complex mechanisms of DNA inter strand cross-link repair. Cold Spring Harb Perspect Med 3, 1–25 (2013).
-
- McHugh P. J. XPF–ERCC1: Linchpin of DNA crosslink repair. PLoS Genet 16, 1–5 (2020).
Publication types
LinkOut - more resources
Full Text Sources
