

doi: 10.1038/311721a0.
Resolution of synthetic att-site Holliday structures by the integrase protein of bacteriophage lambda
- PMID: 6092961
- PMCID: PMC1965284
- DOI: 10.1038/311721a0
Item in Clipboard
Resolution of synthetic att-site Holliday structures by the integrase protein of bacteriophage lambda
Nature.
1984 Oct.
Abstract
Site-specific recombination of the bacteriophage lambda genome into and out of the host bacterial genome is postulated to involve the formation of Holliday structure intermediates by reciprocal single-strand exchanges. Synthetic analogues of the predicted recombination intermediates are resolved in vitro by the protein product of the lambda int gene. Some of the structural features and reaction conditions for this genetic recombination can now be defined.
Figures
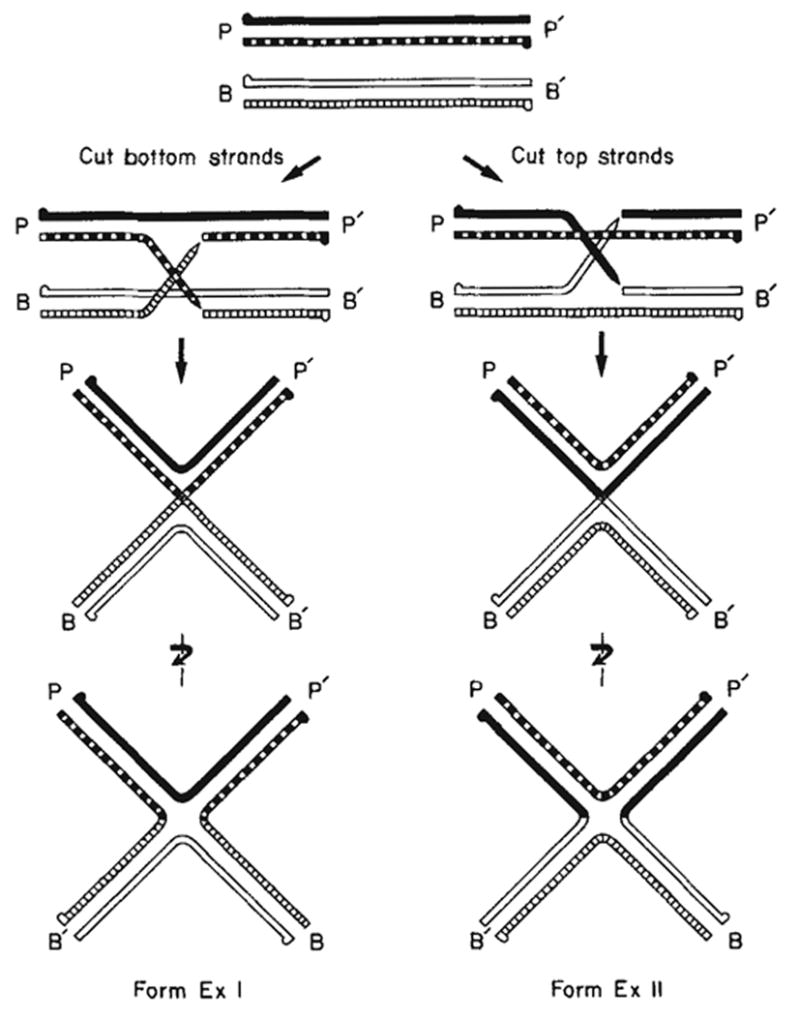
Formation of Holliday structures (χ-forms) during λ site-specific recombination. The attP top (
 ) and bottom (
) and bottom (
 ) strands and the attB top (
) strands and the attB top (
 ) and bottom (
) and bottom (
 ) strands have their 5′ termini indicated by a knob. The cutting and exchange of two bottom strands (left) results in form ExI, whereas the cutting and exchange of two top strands (right) results in form ExII. In the case where attL and attR are the parental DNAs (not shown here), the cutting and exchange of the two top strands would result in form ExI and the cutting and exchange of the two bottom strands would result in form ExII.
) strands have their 5′ termini indicated by a knob. The cutting and exchange of two bottom strands (left) results in form ExI, whereas the cutting and exchange of two top strands (right) results in form ExII. In the case where attL and attR are the parental DNAs (not shown here), the cutting and exchange of the two top strands would result in form ExI and the cutting and exchange of the two bottom strands would result in form ExII.
) and bottom (
) strands and the attB top (
) and bottom (
) strands have their 5′ termini indicated by a knob. The cutting and exchange of two bottom strands (left) results in form ExI, whereas the cutting and exchange of two top strands (right) results in form ExII. In the case where attL and attR are the parental DNAs (not shown here), the cutting and exchange of the two top strands would result in form ExI and the cutting and exchange of the two bottom strands would result in form ExII.
Construction of forms ExI and ExII Holliday structures with a branch point in the common core region of the att site. Equivalent amounts of the four parental fragments, containing the POP’, BOP’, POB’ and BOB’ DNA sequences, were denatured and reannealed as described here, a, Schematic illustration of the stepwise formation of χ-structures during the reannealing process. Designations for the attP top (
) and bottom (
) strands and the attB top (
) and bottom (
) strands are also used to indicate the corresponding DNA strands in the prophage att sites BOP’ and POB’. The 5′ termini are marked with a knob. b, The ethidium bromide-stained agarose gel shows the four DNA restriction fragments before (lane 1) and after (lane 2) denaturation and after reannealing (lane 3). Electron micrograph shows DNA eluted from the gel band marked χ-form. Methods: 40-120 pmol of restriction fragments in 0.8-1.0 ml were denatured and reannealed by one of the following two procedures: (1), the DNAs were denatured by boiling at 100 °C for 2 min in 1 mM Tris-HCl (pH 7.9), 0.1 mM EDTA, followed by rapid chilling in ice water. One-tenth volume of 20 × SSC (3 M NaCl, 0.3 M Na citrate) was added and the mixture was placed in a 65 °C water bath that was then allowed to equilibrate to room temperature overnight. (2), DNA restriction fragments in low melting agarose were heated at 68 °C until the agarose was melted and then placed at 37 °C. They were then denatured in 0.1 M NaOH for 5 min; after adding ½ vol of deionized formamide and neutralizing with acetic acid, they were incubated at 37 °C overnight. The χ-forms were purified by gel electrophoresis in 1% agarose at 1.8 V cm−1 in E* buffer (18 mM NaCl, 2mM EDTA, 40 mM Tris, 20mM sodium acetate, adjusted to pH 8.0 with acetic acid). DNA was electroeluted from the excised gel band and purified by elution from a BND cellulose column as described previously.
) and bottom (
) strands and the attB top (
) and bottom (
) strands are also used to indicate the corresponding DNA strands in the prophage att sites BOP’ and POB’. The 5′ termini are marked with a knob. b, The ethidium bromide-stained agarose gel shows the four DNA restriction fragments before (lane 1) and after (lane 2) denaturation and after reannealing (lane 3). Electron micrograph shows DNA eluted from the gel band marked χ-form. Methods: 40-120 pmol of restriction fragments in 0.8-1.0 ml were denatured and reannealed by one of the following two procedures: (1), the DNAs were denatured by boiling at 100 °C for 2 min in 1 mM Tris-HCl (pH 7.9), 0.1 mM EDTA, followed by rapid chilling in ice water. One-tenth volume of 20 × SSC (3 M NaCl, 0.3 M Na citrate) was added and the mixture was placed in a 65 °C water bath that was then allowed to equilibrate to room temperature overnight. (2), DNA restriction fragments in low melting agarose were heated at 68 °C until the agarose was melted and then placed at 37 °C. They were then denatured in 0.1 M NaOH for 5 min; after adding ½ vol of deionized formamide and neutralizing with acetic acid, they were incubated at 37 °C overnight. The χ-forms were purified by gel electrophoresis in 1% agarose at 1.8 V cm−1 in E* buffer (18 mM NaCl, 2mM EDTA, 40 mM Tris, 20mM sodium acetate, adjusted to pH 8.0 with acetic acid). DNA was electroeluted from the excised gel band and purified by elution from a BND cellulose column as described previously.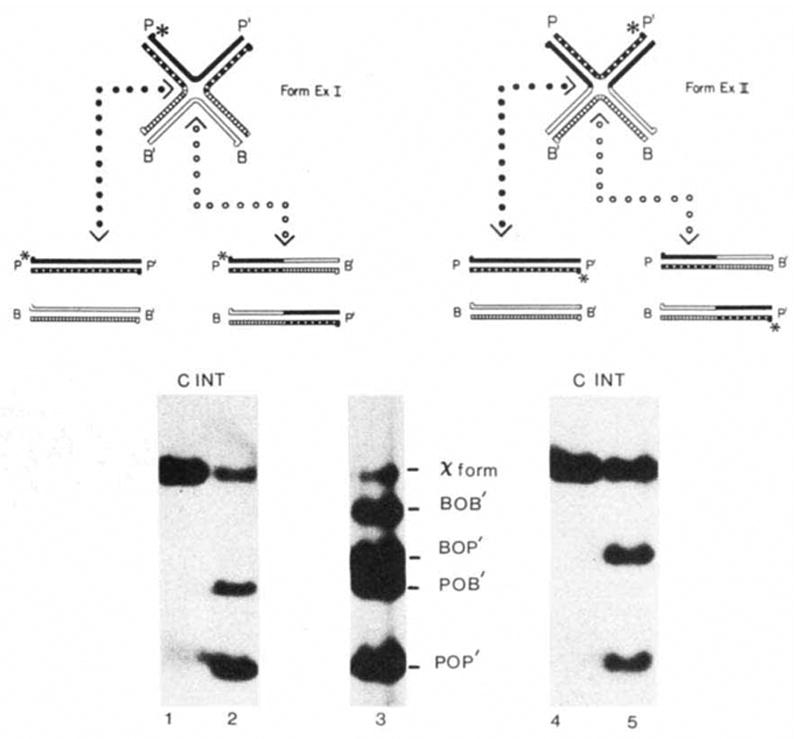
Resolution of forms ExI and ExII by Int protein. The strands of the four att sites and the 5 ′ termini are designated as in Fig. 2. The positions of the unique 32P label (*) in each χ-form that are expected after resolution are shown in the schematic diagram (see text). Lane 3 shows the positions of the χ-forms and the four restriction fragments used to make the χ-forms. Each labelled χ-form was incubated without (C) or with (INT) Int protein. Radioactive bands were visualized by autoradiography. Methods: χ-Forms were constructed according to the first procedure described in Fig. 2 legend using the POP’ HmdIII-BamHI-492 fragment of pWR1, the BOB’ EcoRI-BamHI-1740 fragment of pWR101, the BOP’ EcoRI-BamHI-1210 fragment of pPH201 and the POB’ HindIII-BamHI-1020 fragment of pPH202 (Table 1). The POP’ restriction fragment was labelled before denaturation with [γ-32P]ATP and polynucleotide kinase at the HindIII site (for labelling form ExI) or at the Bam HI site (for labelling form ExII). Each reaction mixture (20 μl) contained 0.1 μg of 32P-labeIled form ExI or form ExII in 50 mM Tris-HCl (pH 7.9), 5 mM EDTA, 50 mM KC1 and 6.25 mM spermidine. Incubation with 1 U of Int protein (purified by N. Hasan) was carried out at 25 °C for 60 min. (In these reaction conditions 1 U of Int plus 1 U of purified IHF, gives optimal recombination of 0.1 pmol supercoiled attP, pWRl, and 0.1 pmol of linear attB, pWR101.) The reactions were terminated with SDS (0.1% final) and electrophoresed on a 1% agarose gel as described for Fig. 2.

Construction of a non-att Holliday structure. The HindIII fragment of λ DNA (
 ), which extends from 57.0% to 52.4% on the λ map and does not contain an att site, was cloned as a tandem duplication into the HindIII site of pBR322 (—), to generate plasmid pPH910. Three HinfI fragments of pPH910 were isolated for the χ-form construction: fragment II consists of the left pBR322-λ junction, fragment III of the head-to-tail junction of the two λ fragments, and fragment IV of the right λ-pBR322 junction. Fragment I is the pBR322 HinfI fragment into which the λ DNA had been inserted. For each fragment, and for each arm of the χ-structure, the amount (base pairs) of pBR322 DNA or λ DNA is indicated. Methods: The χ-form was generated by alkali denaturation and renaturation according to scheme (2) and purified by agarose gel electrophoresis as described in Fig. 2 legend.
), which extends from 57.0% to 52.4% on the λ map and does not contain an att site, was cloned as a tandem duplication into the HindIII site of pBR322 (—), to generate plasmid pPH910. Three HinfI fragments of pPH910 were isolated for the χ-form construction: fragment II consists of the left pBR322-λ junction, fragment III of the head-to-tail junction of the two λ fragments, and fragment IV of the right λ-pBR322 junction. Fragment I is the pBR322 HinfI fragment into which the λ DNA had been inserted. For each fragment, and for each arm of the χ-structure, the amount (base pairs) of pBR322 DNA or λ DNA is indicated. Methods: The χ-form was generated by alkali denaturation and renaturation according to scheme (2) and purified by agarose gel electrophoresis as described in Fig. 2 legend.
), which extends from 57.0% to 52.4% on the λ map and does not contain an att site, was cloned as a tandem duplication into the HindIII site of pBR322 (—), to generate plasmid pPH910. Three HinfI fragments of pPH910 were isolated for the χ-form construction: fragment II consists of the left pBR322-λ junction, fragment III of the head-to-tail junction of the two λ fragments, and fragment IV of the right λ-pBR322 junction. Fragment I is the pBR322 HinfI fragment into which the λ DNA had been inserted. For each fragment, and for each arm of the χ-structure, the amount (base pairs) of pBR322 DNA or λ DNA is indicated. Methods: The χ-form was generated by alkali denaturation and renaturation according to scheme (2) and purified by agarose gel electrophoresis as described in Fig. 2 legend.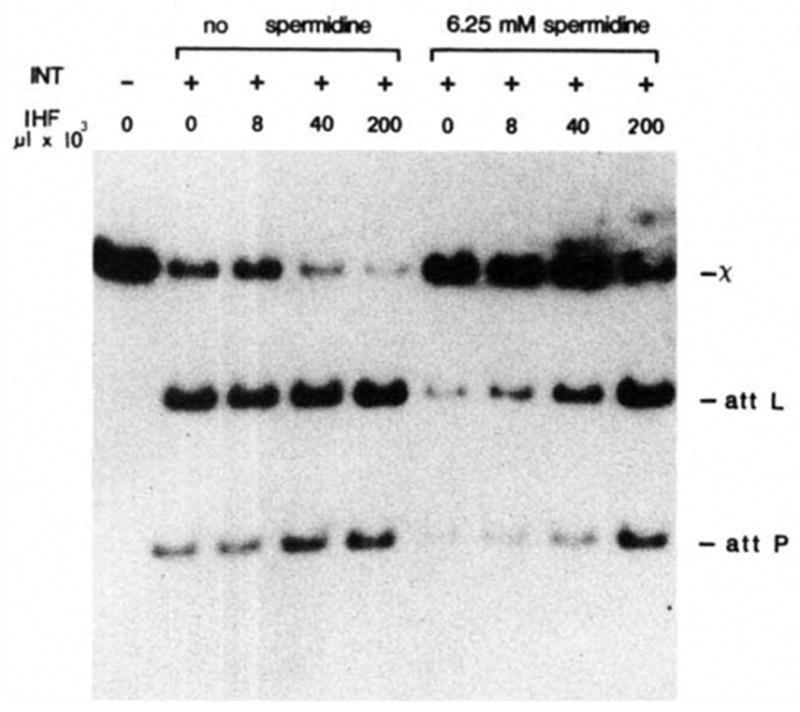
Effect of IHF protein and spermidine on resolution. 32P-labelled form ExII was constructed and isolated as described in Fig. 3 legend. The composition and processing of each resolution reaction was the same as described for Fig. 3 except that incubation with 0.2 U of purified Int protein was for 30 min. Purified IHF protein (a gift from H. Nash) at 1 U μl −1 and spermidine were present as indicated.

Int resolution of χ-forms lacking one or more arm-type Int binding sites. χ-forms composed of intact att sites (W.T.) or lacking the indicated arm-type Int binding site(s) (−P1, P2, etc.) were constructed from the appropriate HincII restriction fragments (Table 1, using the second procedure in Fig. 2). b, The attP coordinates for the 15 bp common core (
 ); the consensus recognition sequences (
); the consensus recognition sequences (
 ) and the approximate region of DNA covered (as determined by nuclease protection) for each of the following: the junction-type Int binding sites, C and C’ (
) and the approximate region of DNA covered (as determined by nuclease protection) for each of the following: the junction-type Int binding sites, C and C’ (
 ), the arm-type Int binding sites P1, P2, P’1, P’2, and P’3 (
), the arm-type Int binding sites P1, P2, P’1, P’2, and P’3 (
 ),, the IHF binding sites H1, H2 and H’ (
),, the IHF binding sites H1, H2 and H’ (
 ). The endpoints of att DNA in the three deletions used to construct the different χ-structures are indicated (−109, −89 and +46) (see Table 1). Each χ-form was incubated in the absence (−) or presence (+) of purified Int protein in a 20-μl reaction that contained ~0.01 μg of χ-forrn, 0.5 μg of supercoiled pBR322 DNA and 1 U of purified Int. After electrophoresis of the reaction products, gels were stained with ethidium bromide (0.5 μg ml−1) to visualize the nicking-closing activity of Int on the supercoiled pBR322 DNA. The expected levels of activity were observed in each case (data not shown). The 32P-labelled χ-forms and resolution products were visualized by autoradiography. The size (in base pairs) of each labelled resolution product is as follows: BB’, 940; BP’, 1,077; BΔ’, 886; PB’, 1,114; ΔB’, 972 (lane 4) and 952 (lanes 6 and 8). In this exposure of the autoradiogram, resolution products of similar size are not clearly resolved in several of the lanes; shorter exposures (and other gels) show that both resolution products are obtained in each reaction. The incomplete resolution observed in lane 6 of the upper gel is not typical; in other experiments in these conditions resolution is complete. Methods: Reaction conditions, processing and gel electrophoresis were as for Fig. 3, with no spermidine. For 32P labelling of forms ExI and ExII, the BOB’ HincII fragment was first labelled at the 5′ termini with polynucleotide kinase and then strand-separated by gel electrophoresis. The top strand of BOB’ was incorporated into form ExI and the bottom strand into form ExII (see Fig. 2) by annealing with the other three restriction fragments (Table 1). The specific activity is the same for the entire form ExI family (or the entire form ExII family), because the 32P label is always on the BOB’ strand which is common to all five χ-forms (see Table 1).
). The endpoints of att DNA in the three deletions used to construct the different χ-structures are indicated (−109, −89 and +46) (see Table 1). Each χ-form was incubated in the absence (−) or presence (+) of purified Int protein in a 20-μl reaction that contained ~0.01 μg of χ-forrn, 0.5 μg of supercoiled pBR322 DNA and 1 U of purified Int. After electrophoresis of the reaction products, gels were stained with ethidium bromide (0.5 μg ml−1) to visualize the nicking-closing activity of Int on the supercoiled pBR322 DNA. The expected levels of activity were observed in each case (data not shown). The 32P-labelled χ-forms and resolution products were visualized by autoradiography. The size (in base pairs) of each labelled resolution product is as follows: BB’, 940; BP’, 1,077; BΔ’, 886; PB’, 1,114; ΔB’, 972 (lane 4) and 952 (lanes 6 and 8). In this exposure of the autoradiogram, resolution products of similar size are not clearly resolved in several of the lanes; shorter exposures (and other gels) show that both resolution products are obtained in each reaction. The incomplete resolution observed in lane 6 of the upper gel is not typical; in other experiments in these conditions resolution is complete. Methods: Reaction conditions, processing and gel electrophoresis were as for Fig. 3, with no spermidine. For 32P labelling of forms ExI and ExII, the BOB’ HincII fragment was first labelled at the 5′ termini with polynucleotide kinase and then strand-separated by gel electrophoresis. The top strand of BOB’ was incorporated into form ExI and the bottom strand into form ExII (see Fig. 2) by annealing with the other three restriction fragments (Table 1). The specific activity is the same for the entire form ExI family (or the entire form ExII family), because the 32P label is always on the BOB’ strand which is common to all five χ-forms (see Table 1).
); the consensus recognition sequences (
) and the approximate region of DNA covered (as determined by nuclease protection) for each of the following: the junction-type Int binding sites, C and C’ (
), the arm-type Int binding sites P1, P2, P’1, P’2, and P’3 (
),, the IHF binding sites H1, H2 and H’ (
). The endpoints of att DNA in the three deletions used to construct the different χ-structures are indicated (−109, −89 and +46) (see Table 1). Each χ-form was incubated in the absence (−) or presence (+) of purified Int protein in a 20-μl reaction that contained ~0.01 μg of χ-forrn, 0.5 μg of supercoiled pBR322 DNA and 1 U of purified Int. After electrophoresis of the reaction products, gels were stained with ethidium bromide (0.5 μg ml−1) to visualize the nicking-closing activity of Int on the supercoiled pBR322 DNA. The expected levels of activity were observed in each case (data not shown). The 32P-labelled χ-forms and resolution products were visualized by autoradiography. The size (in base pairs) of each labelled resolution product is as follows: BB’, 940; BP’, 1,077; BΔ’, 886; PB’, 1,114; ΔB’, 972 (lane 4) and 952 (lanes 6 and 8). In this exposure of the autoradiogram, resolution products of similar size are not clearly resolved in several of the lanes; shorter exposures (and other gels) show that both resolution products are obtained in each reaction. The incomplete resolution observed in lane 6 of the upper gel is not typical; in other experiments in these conditions resolution is complete. Methods: Reaction conditions, processing and gel electrophoresis were as for Fig. 3, with no spermidine. For 32P labelling of forms ExI and ExII, the BOB’ HincII fragment was first labelled at the 5′ termini with polynucleotide kinase and then strand-separated by gel electrophoresis. The top strand of BOB’ was incorporated into form ExI and the bottom strand into form ExII (see Fig. 2) by annealing with the other three restriction fragments (Table 1). The specific activity is the same for the entire form ExI family (or the entire form ExII family), because the 32P label is always on the BOB’ strand which is common to all five χ-forms (see Table 1).References
-
- Nash HAA. Rev Genet. 1981;15:143–167. - PubMed
-
- Weisberg R, Landy A. In: Lambda II. Hendrix RW, Roberts JW, Stahl FW, Weisberg RW, editors. Cold Spring Harbor Laboratory; New York: 1983. pp. 211–250.
-
- Nash HA, Robertson CA. J biol Chem. 1981;256:9246–9253. - PubMed
-
- Abremski K, Gottesman S. J biol Chem. 1982;257:9658–9710. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
