

doi: 10.1038/285085a0.
The lambda phage att site: functional limits and interaction with Int protein
- PMID: 6246439
- PMCID: PMC1994824
- DOI: 10.1038/285085a0
Item in Clipboard
The lambda phage att site: functional limits and interaction with Int protein
Nature.
.
Abstract
Site specific integrative recombination of bacteriophage lambda involves unequal partners. The minimal phage att site is composed of approximately 240-base pairs and four distinct binding sites for Int protein, at least three of which are crucial for function. This 'donor site' recombines efficiently with a smaller 'recipient site' that lacks the extensive interactions with Int protein.
Figures
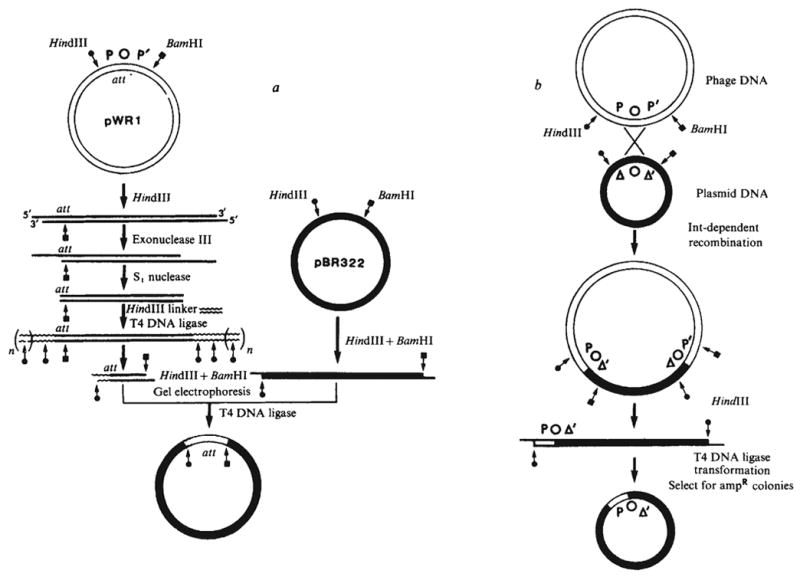
Construction of plasmids carrying resected att sites. a, Plasmid pWR1 was first linearised, for example with HindIII for shortening the P arm. Digestion of the linear DNA with a combination of exonuclease III from E. coli (New England Biolabs) and nuclease S1 (Miles) generates a series of shortened plasmids carrying flush base-paired termini. A chemically synthesised decamer HindIII linker (Collaborative Research) was ligated to the blunt end by T4 DNA ligase. Digestion with HindIII and BamHI generates a family of fragments with unaltered P′ arms and P arms shortened to different extents. These were fractionated by gel electrophoresis and regions corresponding to the desired size were excised and eluted. Finally, the size selected fragments were recloned between the HindIII and BamHI sites of pBR322 and the plasmid carried in each clone characterised by restriction fragment analysis. In an analogous fashion, shortening of the P′ arm would be carried out by using BamHI in the place of HindIII to linearise the plasmid and using a BamHI linker in place of the HindIII linker. b, Cells harbouring a plasmid with a shortened att site arm were infected with λ phage carrying a wild-type att site. As a result, some of the infecting phage underwent Int-dependent recombination with the att site on the plasmid and thus acquired the ability to transduce ampR (D. Kamp and K. Mizuuchi, personal communication). The att site at each phage-plasmid junction has one arm derived from the plasmid and the other arm derived from the phage. Recovery of the recombinant att site on a self-replicating plasmid was accomplished by digesting the plasmid-phage hybrid DNA with HindIII, recircularising the DNA (by means of the HindIII ends) and selecting ampicillin-resistant transformants. These procedures will be described in detail elsewhere (P.-L. H. and A.L., in preparation).
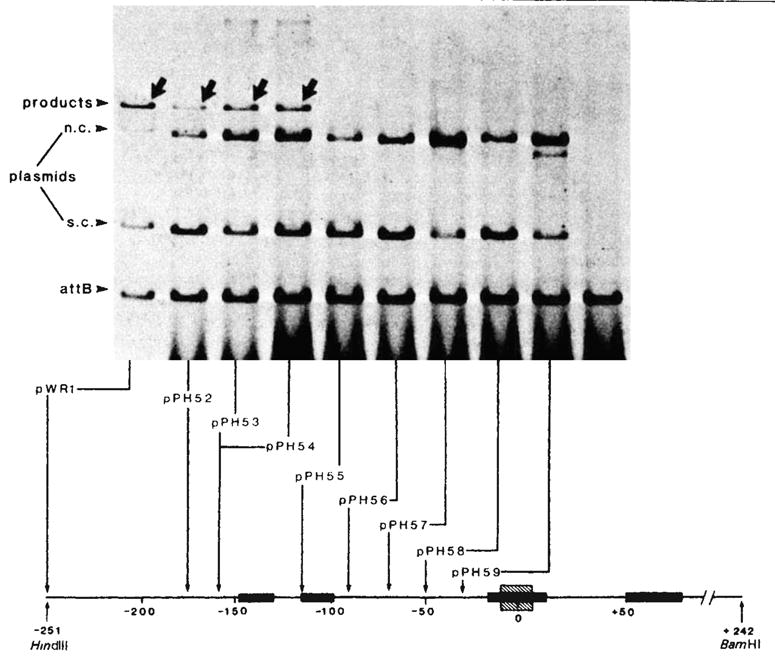
Integrative recombination of plasmids with resected P arms. Each reaction mixture contained ~1 μg supercoiled plasmid DNA with an intact, or P arm-resected, phage att site and 1μg of linear DNA with a bacterial att site (EcoRI-BamHI fragment; 1,600-base pairs) at a molar ratio of 1:3. The recombination was carried out in a 20-μl mixture containing 50 mM Tris-HCl (pH 7.4), 5 mM EDTA, 40 mM KCl, 12.5 mM spermidine, 4 μl of sonicated bacterial extract and 1.5 μl of Int traction 1 (refs 8, 10) After incubation at 25 °C for 40 min, 30 μl of a buffer containing 0.3 M NaCl, 10 mM Tris-HCl (pH 7.4) was added and the samples were extracted with phenol-CHCl3 before being loaded onto a 1 % agarose slab gel and electrophoresed at 35 V for 16 h in E buffer (40 mM Tris, 20 mM Na-acetate, 2 mM EDTA, 18 mM NaCl, adjusted to pH 7.9 with acetic acid). The gel was strained with EtBr(1 μg ml−1) for 15 min and then photographed under UV light. For quantitation the fragment containing the bacterial att site was first end-labelled with 32P by T4 polynucleotide kinase. After recombination and gel electrophoresis, portions of the gels corresponding to the unreacted linear DNA and the recombination product were excised and counted individually for 32P radioactivities. The percentage of phage att site molecules that recombined with labelled bacterial att site DNA (present in threefold molar excess) was calculated as follows: The P arm end point of each resected plasmid, as determined by restriction enzyme analysis or by DNA sequencing is shown at the bottom (also see Table 1). All plasmids contain a P′ arm extending to the BamHI site at position + 242. Positions on the electropherogram are indicated for the 32P-labelled linear DNA containing the bacterial att site (attB), the unlabelled circular plasmid DNA containing the intact or resected phage att sites and migrating as supercoils (s.c.) or nicked circles (n.c.), and the linear 32P-labelled products of recombination (arrows). Binding sites for Int protein (■) (see legends to Figs 4 and 5), and the 15-base pair common core sequence (hatched) are indicated on the linear map of the att site region.
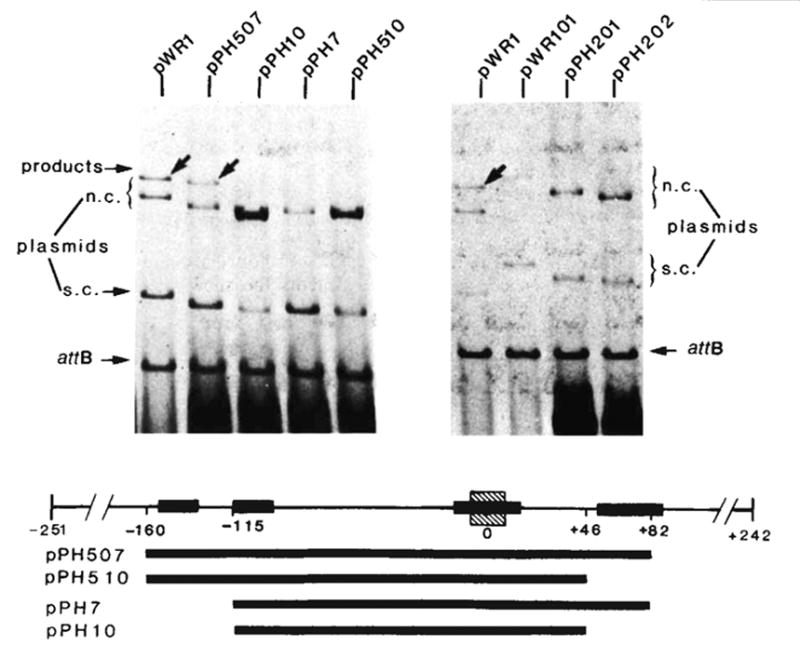
Integrative recombination of the minimal phage att site. The in vitro recombination reaction and gel electrophoresis were carried out as described in Fig. 2 legend. The structure and boundaries of the cloned fragments used in the left panel were confirmed by restriction fragment analysis and by DNA sequencing. The extent of each cloned fragment is indicated by a long bar beneath the map of the att site region. The att sites carried by plasmids in the right panel are: pWR1-phage att site (HindIII-BamHI-492), pWR101-bacterial att site (EcoRI-BamHI-1,600), pPH201-left prophage att site (EcoRI-BamHI-1,300) and pPH202-right prophage att site (HindIII-BamHI-800). Positions on the electropherogram are indicated for the ‘recipient’ linear DNA containing the bacterial att site (attB), the ‘donor’ circular plasmid DNA containing truncated or intact phage att site and migrating as supercoils (s.c.) or nicked circles (n.c.) and the linear products of recombination (arrows). Binding sites for Int protein (■) (see legends to Figs 4 and 5), and the 15-base pair common core sequence (hatched) are indicated on the linear map of the att site region.
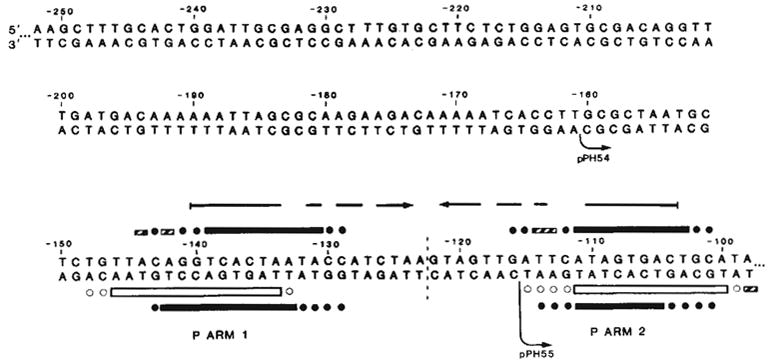
Sequence of the left portion of the P arm and location of Int binding sites P arm 1 and P arm 2. P arm sequence is given negative numbers proceeding leftward from the centre of the common core region (0). We have previously reported the sequence from −114 to +203 (ref. 13). DNA sequence was determined for both strands (with the exception of −252 to −237) by the method of Maxam and Gilbert using 5′ termini labelled with 32P by polynucleotide kinase: top strand sequence from −241 to −90 was obtained from the HindIII end of the HindIII (−251)-Alu(−6) fragment (see ref. 13); top strand sequence from −252 to −241 was determined using the EcoRI site of pBR322 in the plasmid carrying the HindIII(−251)-BamHI(+242) att fragment (pWR1. see text). Complementary sequence from −116 to −237, was obtained from the HinfI end of fragment HinfI(−115)-Mnl 1(recognition site at —231). The left boundaries of the shortest functional P arm (−160, pPH54) and the longest nonfunctional P arm (−115, pPH55) obtained in these experiments are indicated (↪) The locations of Int-binding sites P arm 1 and P arm 2 were determined by the method of DNase I footprinting with modifications described previously (see also Fig. 5). Int protection of sequences in the top strand from neocarzinostatin, digestion (■) was carried out using two different fragments, each labelled at the HindIII end: HindIII(−251)-Alu(−6), carrying only P arm sequences, and HindIII(−251)-HpaII(+305), carrying the entire minimal phage att site sequence (shown in Fig. 5a). Int protection of the bottom strand from neocarzinostatin (■) or DNase I (□) digestion was carried out on a fragment from plasmid pPH310 containing att sequence from +46 to −251 (HindIII), labelled at the BamHI linker adjacent to position +46. Partially protected bases are designated (
 ). Ambiguities in the precise boundaries of the protected regions are due to the failure of neocarzinostatin (●) or DNase I (○) to cut certain bases in the control digests. An inverted repeat structure (→ ←) which includes sequence from each of the two Int-binding sites plus additional sequence between the two sites is indicated.
). Ambiguities in the precise boundaries of the protected regions are due to the failure of neocarzinostatin (●) or DNase I (○) to cut certain bases in the control digests. An inverted repeat structure (→ ←) which includes sequence from each of the two Int-binding sites plus additional sequence between the two sites is indicated.
). Ambiguities in the precise boundaries of the protected regions are due to the failure of neocarzinostatin (●) or DNase I (○) to cut certain bases in the control digests. An inverted repeat structure (→ ←) which includes sequence from each of the two Int-binding sites plus additional sequence between the two sites is indicated.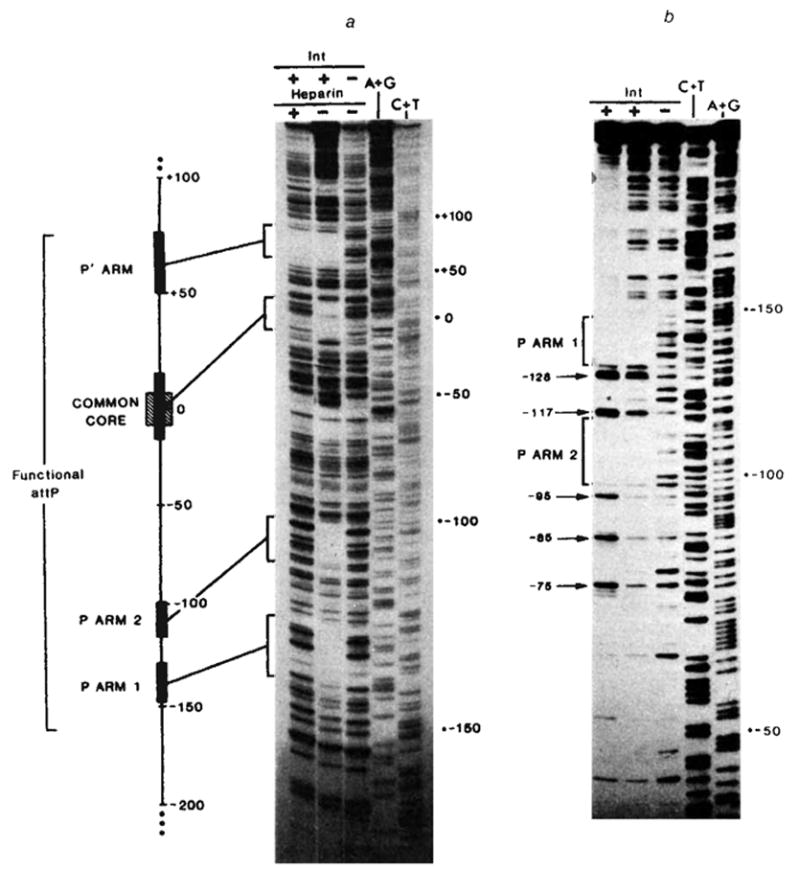
Footprints of Int binding on restriction fragments containing: a, the entire minimal phage att site (using neocarzinostatin); or b, the P arm and common core regions (using DNase I), a, In lanes 1–3 the restriction fragment HindIII(−251)-HpaII(+305), 32P-labeIled at the 5′ end of the top strand (HindIII), was partially digested with neocarzinostatin, in the absence (lane 3) or presence of purified Int protein at 10 μg ml−1 (lane 2) or 20 μgml−1 (lane 1). The Int-DNA complex in lane 1 was challenged with heparin before neocarzinostatin digestion. Sequence markers for this restriction fragment, A+G (lane 4), and C+T (lane 5), were prepared according to the method of Maxam and Gilbert. Details of these methods and electrophoresis conditions have been described previously. A linear map of the minimal phage att site region depicts the relative sizes and positions of the four Int protected sequences (■) seen in the neocarzinostatin footprint. Also indicated are the boundaries (−160 and +82) of the smallest functional attP region obtained in these experiments (see Fig. 3) and the 15-base pair common core sequence (hatched), b, In lanes 1–3 a restriction fragment from plasmid pPH310, containing att sequence from +46 to −251(HindIII) (labelled at the 5′ end of the bottom strand at the BamHI linker adjacent to position +46), was partially digested with DNase I in the absence (lane 3) or presence of purified Int protein at 2.5 μg ml−1 (lane 2) or 5 μg ml−1 (lane 1). The footprinting and electrophoresis conditions were as described previously. The common core region has been run off this gel. Positions of enhanced cutting by DNase I (→) at ~ 10-base pair intervals (−75, −85, −95, −117, −128) are indicated in lane 1.
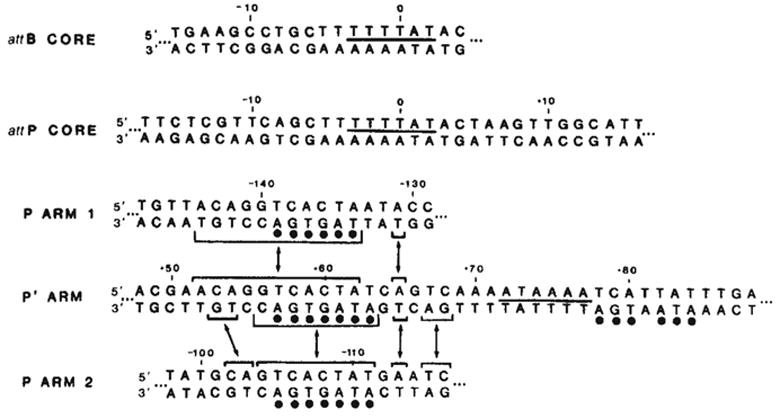
Comparison of Int-protected sequences in the bacterial and phage att sites. The sequence shown for each binding site is the maximum length of DNA protected by Int from either neocarzinostatin or DNase I digestion (see Figs 4 and 5 and ref. 9). The sequence of the P arm 2 site has been inverted (compare Fig. 4) to facilitate comparison of sequence homologies. A 6-base pair homology between the two common core sites and the P′ arm site is indicated by a line between the two strands of sequence (—). Homologies between the left portion of the P′ arm site and each of the two P arm sites are indicated(–) (see text). A 7-base pair sequence in the left half of the P′ arm protected region, found within the sequence shared with the two P arm sites, is also found, with one mismatch, in the right half of the P′ arm site (●).
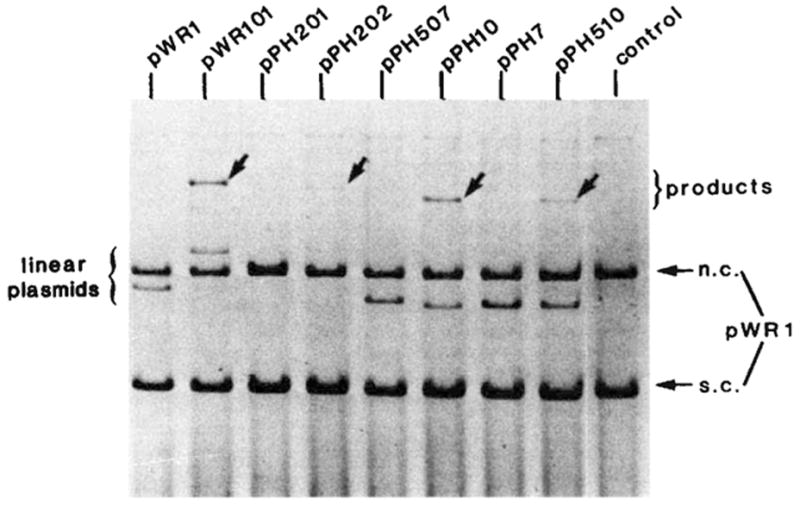
The efficiency of different att sites as linear recombination partners. The in vitro recombination reactions and gel electrophoresis were performed as described in Fig. 2 legend. Each reaction mixture contained supercoiled pWR1 (1.5μg) and linear recipient DNA in a three to one molar ratio. The linear plasmids were generated by PstI digestion of pPH7, 10, 507, 510 or by EcoRI digestion of pWR1, pWR101, pPH201 and pPH202. The origin and boundaries of the restriction fragments carried by each plasmid are described in Fig. 3 legend. Positions are indicated on the electropherogram for DNA containing different ‘recipient’ att sites (linear plasmid), the ‘donor’ circular plasmid DNA (pWR1) containing the phage att site and migrating as supercoils (s.c.) or nicked circles (n.c.) and the linear product of recombination (arrows).
References
-
- Gottesman ME, Weisberg RA. In: The Bacteriophage Lambda. Hershey AD, editor. Cold Spring Harbor Laboratory; New York: 1971. pp. 113–138.
-
- Nash HA. Curr Topics Microbiol Immun. 1978;78:171–199. - PubMed
-
- Miller HI, Kikuchi A, Nash HA, Weisberg RA, Friedman DI. Cold Spring Harb Symp quant Biol. 1979;43:1121–1126. - PubMed
-
- Gottesman S, Abremski K. J molec Biol. 138 (in the press)
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
