

doi: 10.1038/312509a0.
Mechanism of colour discrimination by a bacterial sensory rhodopsin
- PMID: 6504161
- PMCID: PMC4996264
- DOI: 10.1038/312509a0
Item in Clipboard
Mechanism of colour discrimination by a bacterial sensory rhodopsin
Nature.
1984 Dec.
Abstract
A photosensitive protein resembling the visual pigments of invertebrates enables phototactic archaebacteria to distinguish colour. This protein exists in two spectrally-distinct forms, one of which is a transient photoproduct of the other and each of which undergoes photochemical reactions controlling the cell's swimming behaviour. Activation of a single pigment molecule in the cell is sufficient to signal the flagellar motor. This signal-transduction mechanism makes evident a colour-sensing capability inherent in the retinal/protein chromophore.
Figures
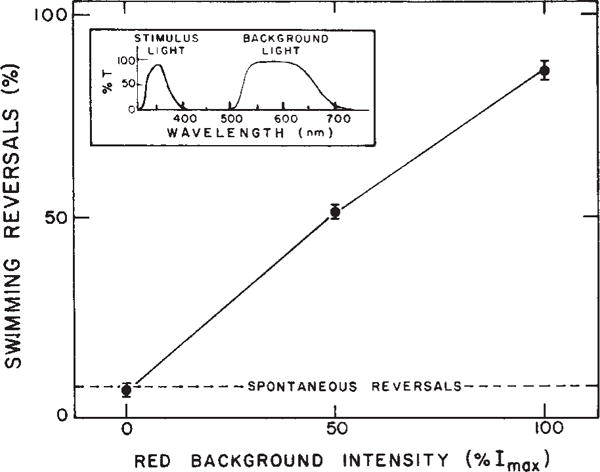
Dependence of near ultraviolet repellent response on visible background light. The figure shows the % of cells reversing within 2 s of the near UV stimulus as a function of red background intensity. Points are average values ± extreme values of 2 separate determinations, each based on >40 cells. The inset shows the transmittance spectra of the filter combinations used for stimulus and visible background irradiation. The stimulus and observation light from a 150 W mercury (Hg) lamp and 50 W tungsten Zeiss microscope illuminator, respectively, were combined through a glass beam splitter which allowed passage of 30% of the infrared observation light (Kodak 87A filter) and 30% of the Hg lamp radiation. The combined beam illuminated the cells through the microscope phase condensor. The red background light was provided by an optically collimated 50 W tungsten light source reflected by a quartz beam-splitter mounted below the condenser. Light fluxes at the sample were measured with a Kettering 65 Radiometer and intensities calculated using irradiation areas measured with a calibrated ocular Zeiss micrometer. Light intensity was varied with Kodak neutral density filters. Stimulus light intensity, 1,400 erg cm−2 s−1. Maximum red background intensity (Imax), 7.2 × 104 erg cm−2 s−1. Infrared radiation, >750 nm.
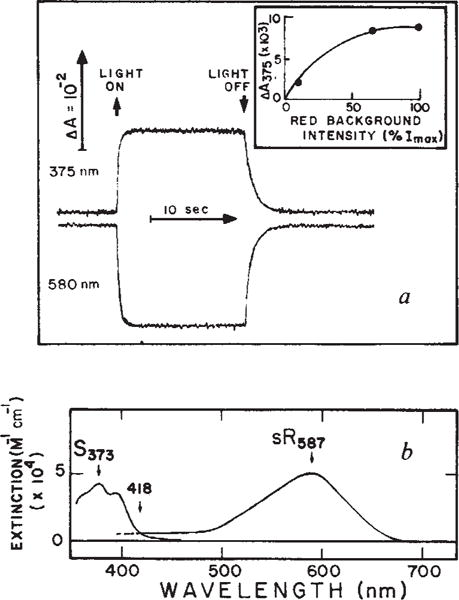
a, Absorbance changes of sensory rhodopsin (sR) at 375 and 580 nm induced by red background light. The sample was a suspension of Flx3R vesicles, prepared vesicles, prepared and regenerated in a 3 × 3 mm quartz cuvette clear on four sides with all-trans retinal as described. The regeneration of sensory rhodopsin was monitored as described, use of the regenerated system permitting precise quantitation of the sensory rhodopsin content. Flx3 vesicles containing native sensory rhodopsin gave the same phototransients as the regenerated protein in Flx3R shown here. Flash-induced absorbance changes were measured as described. Background excitation was provided by a tungsten halogen lamp beam passed through a long pass Corning 2–61 (cutoff 610 nm) and Ditric Optics 700 nm short pass filter combination and applied to the sample at 90° with respect to the measuring beam. The maximum red light intensity (Imax) was 4.25 × 106 erg cm−2 s−1. The inset shows the light intensity-dependence of the absorbance changes at 375 nm (ΔA375). b, Spectrum of S373 and sR587. The latter is the spectrum obtained by regeneration of sensory rhodopsin apoprotein with all-trans retinal and the former was calculated by adding the sR587 spectrum to the light-induced difference spectrum obtained using the system described in a. The extinction coefficients are based on our measurements, and have absolute uncertainty of ±15%.
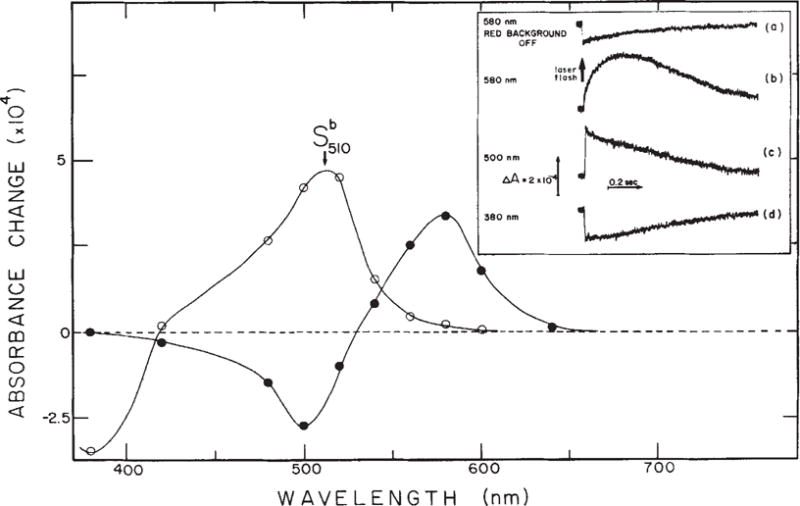
Flash-induced absorbance changes of S373. Inset, transient absorption changes of the sample shown in Fig. 2 in the absence (a) and in the presence (b–d) of 100% Imax red background illumination at three selected wavelengths. Actinic laser flash: 406 nm, 10 ns, 0.4 mJ. Main figure from traces shown in the inset and at other wavelengths the difference in the amplitude of absorbance change between 0 and 100 μs (○) and between 200 ms and 100 μs (●) were calculated and plotted as functions of wavelength. Note that at 580 nm the ratio of the forward to back reaction absorbance change amplitude is the same as the ratio of absolute absorbances of sR587 and S373 at 406 nm (Fig. 2). This indicates that the photochemical quantum yields of the two reactions are nearly equal.
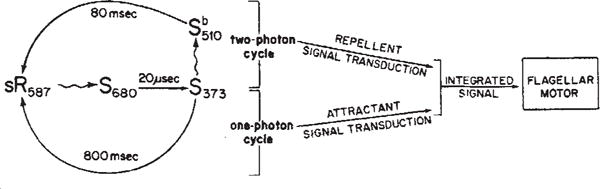
Mechanism of colour discrimination in H. halobium phototaxis. Photon absorption by sR587, the dark-adapted form of sensory rhodopsin, generates a short-lived species S680 which rapidly decays to S373. The S373 form can return to the sR587 form by either of two paths: a purely thermal relaxation or a photoinduced path. If S373 does not absorb a photon, it relaxes thermally to sR587 completing the ‘one-photon’ photocycle. During its transit through the one-photon cycle, the sensory rhodopsin molecule generates an attractant signal to the flagellar motor. If S373 absorbs a photon, it generates the intermediate
. The relaxation of
to sR587 completes a cycle containing two photon absorption steps. After absorption of the second photon in the ‘two-photon’ cycle, sensory rhodopsin generates a repellent signal to the flagellar motor. The attractant and repellent signals are integrated prior to or at their site of action on the flagellar motor.
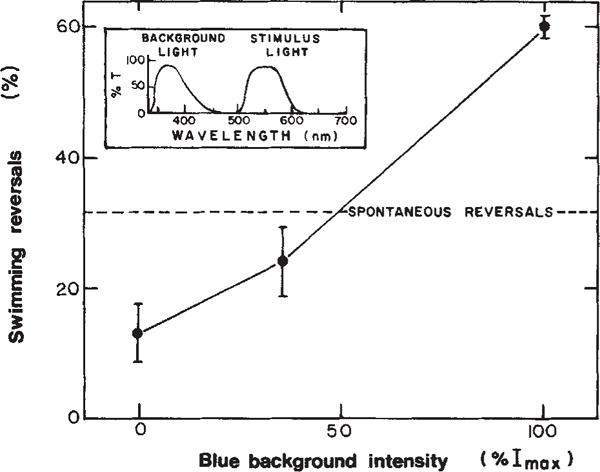
Effect of near ultraviolet background light on cell behavioural response to red light. Experimental conditions as described in Fig. 1 except that the response was assayed as the % reversals within 3 s of a red light stimulus. The range of spontaneous reversal frequencies of populations used in these studies was 14% ± 7% reversing within 2 s. All effects of background illumination on behaviour reported here and in Figs 2 and 6 have been confirmed in populations with both high and low spontaneous reversal frequencies.
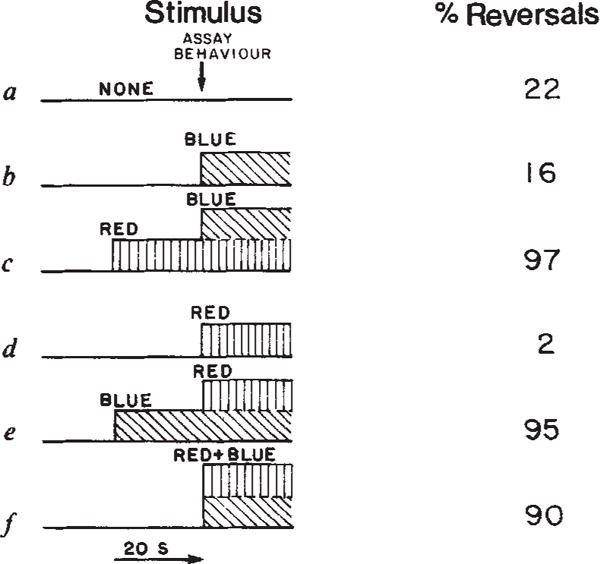
Behavioural response under various permutations of stimulus and background light. Values are averages of duplicate measurements which vary by less than 15%. Experimental conditions as described in Fig. 1 except that reversals were assayed within 3 s of the stimulus, blue and red light were delivered using a tungsten light source and wavelengths were selected by adding or removing filters from the single beam. The Hg light source was blocked in this experiment. ‘Blue’, 380–433 nm (Optics Technology 433 short pass filter+2 mm Plexiglass + two 1–75 Corning infrared absorbing glass), intensity 3.3 × 104 erg cm−2 s−1. ‘Red’, 530–700 nm (Corning 3–68 + two Corning 1–75), intensity 1.0 × 105 erg cm−2 s−1. ‘Red+Blue’, The tungsten beam filtered through two Corning 1–75 + 2 mm Plexiglass, intensity 1.5 × 105 erg cm−2 s−1.
References
-
- Koshland DE., Jr . Bacterial Chemotaxis as a Model Behavioral System. Vol. 2. Raven; New York: 1980. pp. 1–193.
-
- Hildebrand E, Dencher N. Nature. 1975;257:46–48. - PubMed
-
- Spudich JL, Stoeckenius W. J Photobiochem Photobiophys. 1979;1:43–53.
-
- Dencher NA, Hildebrand E. Z Naturforsch. 1979;34:841–847. - PubMed
-
- Sperling W, Schimz A. Biophys Struct Mech. 1980;6:165–169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
