

doi: 10.1038/294422a0.
Dual function transcripts specifying tRNA and mRNA
- PMID: 7312036
- PMCID: PMC1987712
- DOI: 10.1038/294422a0
Item in Clipboard
Dual function transcripts specifying tRNA and mRNA
Nature.
.
Abstract
A cluster of four tRNA genes in Escherichia coli is co-transcribed with an adjacent gene encoding elongation factor Tu. The resultant transcript that specifies both structural (tRNA) and informational (mRNA) RNA may not be an uncommon occurrence and has interesting regulatory implications.
Figures
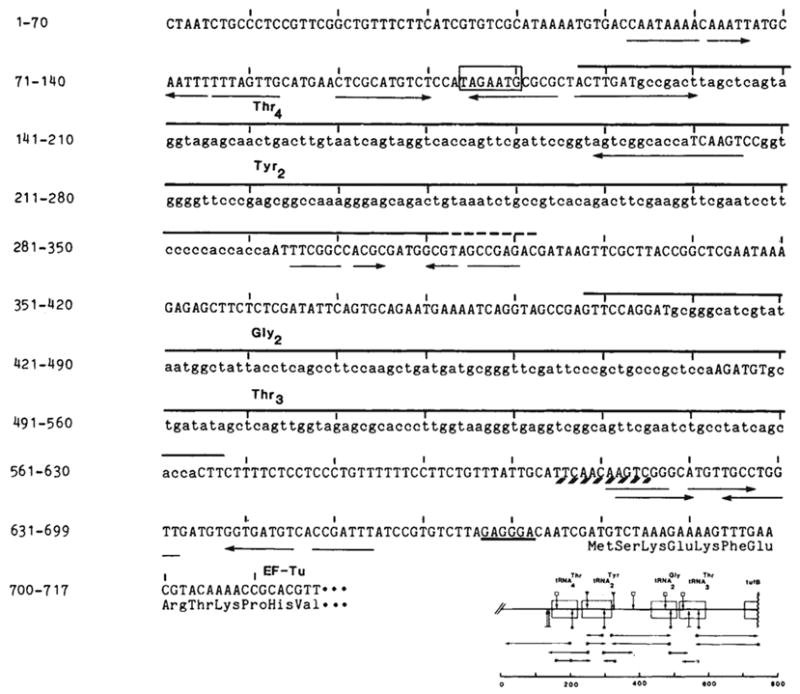
Nucleotide sequences of the tyrU cluster and proximal tufB region. The DNA strand with the sequence of the RNA transcript is shown. The source of DNA was the transducing phage λ rifd 18 (ref. ; supplied by M. Nomura or N. Fiil). DNA sequencing was by the method of Maxam and Gilbert; *, 5′-labelled end of the restriction fragments; →, direction and extent of the sequence obtained; the scale is in base pairs. Complement and overlap were obtained for all new sequences reported here with the exception of the sequence upstream of the
gene. The sequence is identical to that reported by An and Friesen with the exception of an additional base pair they place at position 317. Structural tRNA sequence (lower case) and the two dimeric precursors are overlined. The
immer is 170 bases. The precise 3′ terminus (---) of
(~200 bases) is not known. It is located in a region of dyad symmetry (→ ←) as is the site between the tRNA cluster and the tufB gene at which processing or termination occurs (positions 608–618; see text). Transcription from the presumptive promoter would initiate at the A 7 bases upstream of
as inferred from the results of Taylor and Burgess. The Pribnow box (□) is preceded by a −35 sequence (TTGCAT) similar to the consensus sequence TTGACA of Rosenberg and Court and also byan A+T-rich region (88% A-T from −63 to −38). A potential ribosome binding site, located 6 bases upstream of tufB, is underlined. The first 12 amino acids of tubB (ref. 19) are listed below their respective codons.
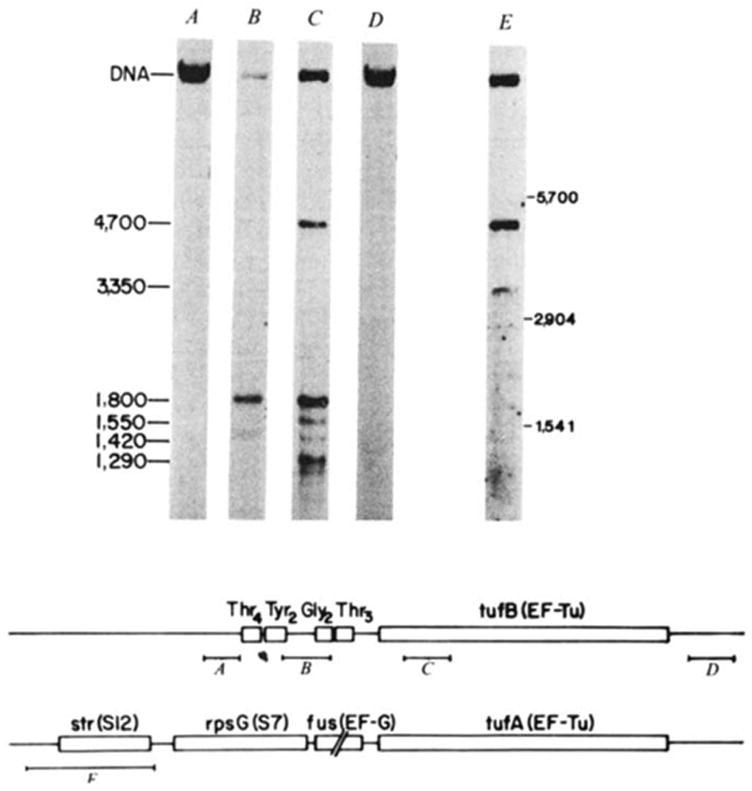
Characterization of the transcript specifying the tyrU cluster and tufB. E. coli strain ABL-1 (RNase III− RNase Pts; ref. ; gift of W. McClain) was grown for several generations at 30 °C, and then incubated at 42 °C for 30 min to inactivate RNase P. RNA was extracted according to Gegenheimer and Apirion as detailed elsewhere, denatured with glyoxal, electrophoresed in 1.5% agarose gels with recirculation of the 10 mM sodium phosphate, pH 6.5, 1 mM EDTA running buffer, and transferred to DBM paper. The RNA blots were probed with DNA fragments (A-E) which were nick translated with [α-32P]dCTP (NEN; 2–3,000 Ci mmol−1) to a specific activity of ~5 × 107 c.p.m. per μg. Hybridizations were carried out in the presence of 10% dextran sulphate as detailed by Alwine et al. but omitting carrier DNA. Blots were washed first in 0.3 M NaCl, 0.03 M sodium citrate, 0.1% SDS at 23 °C for 20 min, and then in 15 mM NaCl, 1.5 mM sodium citrate, 0.1% SDS for 45 min at 50–55 °C. The 32P-labelled probes isolated from the region of λ rifd18 depicted in the diagram were: A, HhaI-145; B, HinfI-190; C, HincII-SmaI-187; D, BstNI-HaeIII-185. Probe E is a HaeIII-530 fragment encompassing the S12 ribosomal protein gene, which was isolated from λ fus3 (obtained from J. Yates and M. Nomura). Molecular weight markers were 16S (1,541 bases), 23S (2,904 bases) and 30S RNA (~5,700 bases). Lanes B, C and E have ~2 μg RNA per lane, and lanes A and D have 30 μg per lane. The scale is in bases. Structural sequences for tRNAs and tufB are boxed.
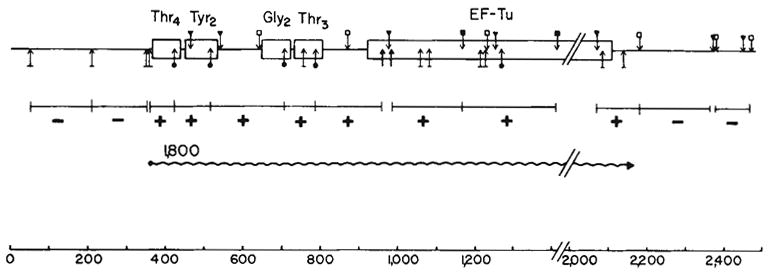
Organization and transcription of the tRNA-tufB region. The locations of the restriction sites were obtained from the sequence shown in Fig. 1 or ref. . Hybridization to the 1,800-base transcript (
 ) is denoted by a + in the appropriate restriction fragment (from λrifd18) and lack of hybridization by −. The HinfI-82 probe from the
region hybridized strongly to 16S RNA in addition to the 1,800-base transcript. This result is consistent with the presence of a number of 12–16-bp homologies between 16S RNA and
sequences. Due to the hybridization with 16S RNA, the HinfI-82 probe could not be used to determine whether the smaller tuf transcripts (1,550 and 1,420 bases shown in Fig. 2) were tufA or tufB specific. The scale is in bases. Structural sequences for the tRNAs and tufB are boxed.
) is denoted by a + in the appropriate restriction fragment (from λrifd18) and lack of hybridization by −. The HinfI-82 probe from the
region hybridized strongly to 16S RNA in addition to the 1,800-base transcript. This result is consistent with the presence of a number of 12–16-bp homologies between 16S RNA and
sequences. Due to the hybridization with 16S RNA, the HinfI-82 probe could not be used to determine whether the smaller tuf transcripts (1,550 and 1,420 bases shown in Fig. 2) were tufA or tufB specific. The scale is in bases. Structural sequences for the tRNAs and tufB are boxed.
 , BstI; ↓, HaeIII; ↥, HhaI;
, BstI; ↓, HaeIII; ↥, HhaI;
 HincII;
HincII;
 HinfI;
HinfI;
 SmaI.
SmaI.
) is denoted by a + in the appropriate restriction fragment (from λrifd18) and lack of hybridization by −. The HinfI-82 probe from the
region hybridized strongly to 16S RNA in addition to the 1,800-base transcript. This result is consistent with the presence of a number of 12–16-bp homologies between 16S RNA and
sequences. Due to the hybridization with 16S RNA, the HinfI-82 probe could not be used to determine whether the smaller tuf transcripts (1,550 and 1,420 bases shown in Fig. 2) were tufA or tufB specific. The scale is in bases. Structural sequences for the tRNAs and tufB are boxed.
, BstI; ↓, HaeIII; ↥, HhaI;
HincII;
HinfI;
SmaI.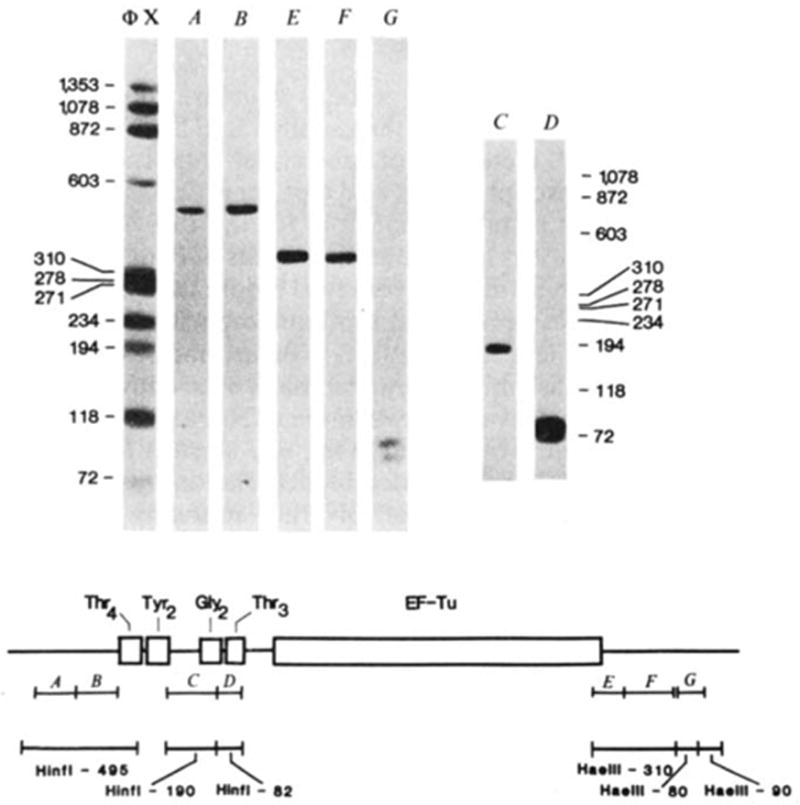
Hybridization of probes to λ rifd18 DNA blotted onto DBM paper. The BamHI-EcoRI-5,500 fragment containing the entire tRNA-tufB region was digested with either HinfI (lanes A–D) or HaeIII (lanes E–G) and 0.1 μg of each digest per slot was electrophoresed on a composite gel (5% acrylamide, 0.7% agarose) cross-linked with N, N′-diallyltartardiamide as detailed by Alwine et al.. The gel was soaked in 2% periodic acid to dissolve the cross-links before blotting. Blots were hybridized to the nick-translated probes illustrated in the diagram and washed as described in Fig. 2 legend: A, HhaI-150; B, HhaI-145; C, HinfI-190; D, HinfI-82; E, HaeIII-BstNI-125; F, BstNI-HaeIII-185; G, BstNI-103. Probes A and B both hybridized only to the HinfI-495 fragment (depicted below the probe) from which they can be isolated by HhaI digestion. Similarly, probes E and F each hybridized to the HaeIII-310 fragment from which they can be isolated by BstNI digestion. The two structural tRNA probes, C and D, hybridized only to their respective HinfI-190 or HinfI-82 bands. Importantly, the
-specific probe (C) did not hybridize to the
fragment with which it has limited homology. Probe G hybridized to two HaeIII bands ~80 and 90 bp, consistent with the existence of one HaeIII site within the BstNI-103 fragment and a HaeIII site closely flanking each end (see Fig. 3). No hybridization was observed to additional bands when higher concentrations of DNA (0.5 μg per slot) or longer exposures were used. On the basis of the exposure times and DNA concentrations used in Fig. 3, the nick-translated probes can be expected to detect as little as 0.1 ng RNA. All the fragments (118–1,353 bp) of the ΦX174 HaeIII digest (molecular weight marker) exhibited comparable band intensities when probed with 32P-end-labelled ΦX174.
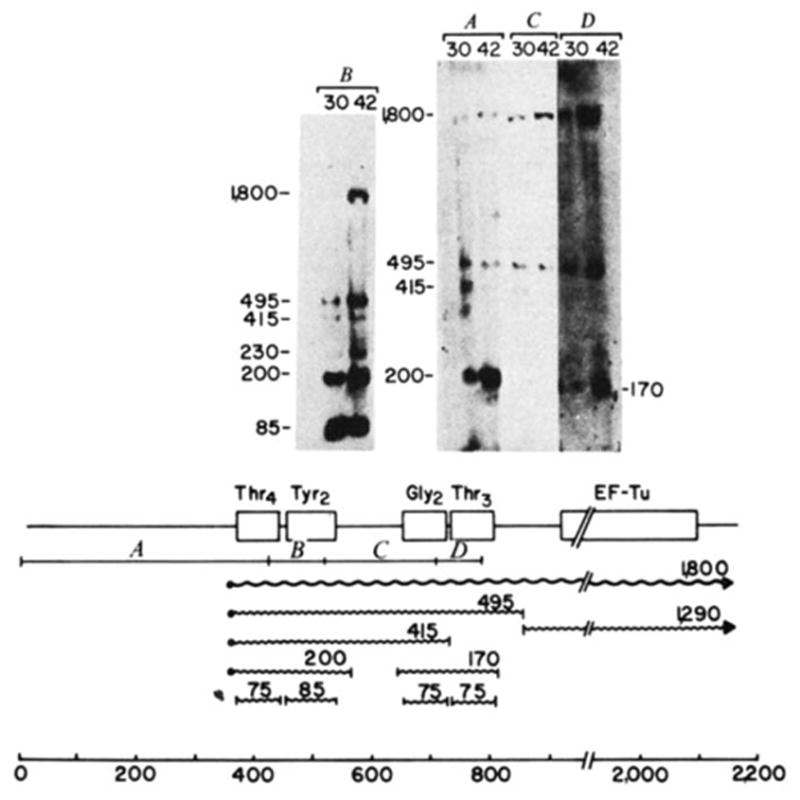
Processing intermediates of the tyrU cluster. RNA was extracted from E. coli strain A49 (RNase Pts; ref. ; gift of W. McClain) grown at either 30 or 42 °C, electrophoresed on 2% agarose gels at 30 μg RNA per slot and blotted onto DBM paper. Transfer conditions were similar to those described for Fig. 2 except that glyoxal was removed with a lower concentration of sodium hydroxide (10 mM compared with 50 mM for Fig. 2). Molecular weight markers were 5′-end-labelled HaeIII and HincII fragments of ΦX174. The gel shown in B was electrophoresed for a shorter time to retain mature tRNA. The 32P-labeIled probes isolated from the regions of λ rifd18 illustrated in the diagram were: A, HinfI-420; B, HinfI-93; C, HinfI-190; D, HinfI-82. The putative primary transcript (
), processing intermediates and mature tRNAs (
 ) are labelled by their respective sizes (in bases) on denaturing gels. The probe containing the structural tyrosine tRNA sequence (B) hybridized to a 230-base transcript that also hybridized with a
upstream probe and must therefore be a tyrT transcript. Probe C contained only 18 bases of structural tyrosine tRNA sequence, which severely limited its binding to the 200- or 230-base tyrosine transcripts in the stringent washing conditions used in these experiments. In the blot shown, the hybridization of probe C to the 170-base transcript did not photograph; in other blots this probe detectably bound the 170-base transcript. The processing scheme is discussed in the text.
) are labelled by their respective sizes (in bases) on denaturing gels. The probe containing the structural tyrosine tRNA sequence (B) hybridized to a 230-base transcript that also hybridized with a
upstream probe and must therefore be a tyrT transcript. Probe C contained only 18 bases of structural tyrosine tRNA sequence, which severely limited its binding to the 200- or 230-base tyrosine transcripts in the stringent washing conditions used in these experiments. In the blot shown, the hybridization of probe C to the 170-base transcript did not photograph; in other blots this probe detectably bound the 170-base transcript. The processing scheme is discussed in the text.
), processing intermediates and mature tRNAs (
) are labelled by their respective sizes (in bases) on denaturing gels. The probe containing the structural tyrosine tRNA sequence (B) hybridized to a 230-base transcript that also hybridized with a
upstream probe and must therefore be a tyrT transcript. Probe C contained only 18 bases of structural tyrosine tRNA sequence, which severely limited its binding to the 200- or 230-base tyrosine transcripts in the stringent washing conditions used in these experiments. In the blot shown, the hybridization of probe C to the 170-base transcript did not photograph; in other blots this probe detectably bound the 170-base transcript. The processing scheme is discussed in the text.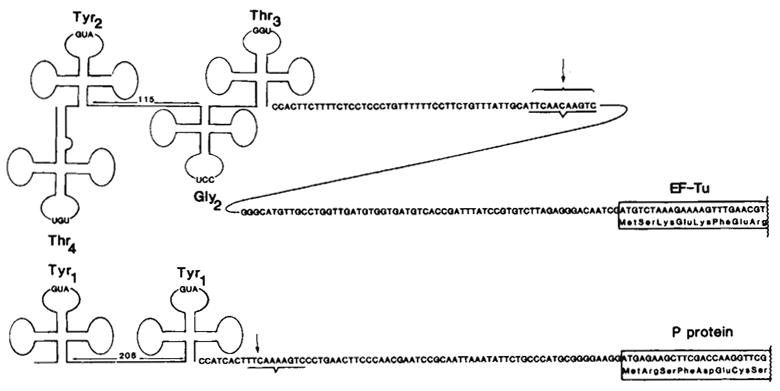
Comparison of the tyrT and tyrU primary transcripts. The tyrT transcript was characterized by Rossi et al. using RNA extracted from an RNase Pts E. coli strain infected with
(doublet). The site conserved between the tyrT and tyrU gene clusters and known to be a 3′-processing site for tyrT transcripts is underlined. For tufB (ref. 19) and for the protamine-like protein (P protein; see text), the first eight amino acids are indicated. Translation of P protein also initiates at the methionine located 4 codons upstream of the start site shown, as indicated by the in vitro production of both a 29- and 33-amino acid basic protein that map to the first repeat.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
