

Review
doi: 10.1152/physrev.1995.75.3.519.
Flow-mediated endothelial mechanotransduction
Affiliations
- PMID: 7624393
- PMCID: PMC3053532
- DOI: 10.1152/physrev.1995.75.3.519
Item in Clipboard
Review
Flow-mediated endothelial mechanotransduction
Physiol Rev.
1995 Jul.
Abstract
Mechanical forces associated with blood flow play important roles in the acute control of vascular tone, the regulation of arterial structure and remodeling, and the localization of atherosclerotic lesions. Major regulation of the blood vessel responses occurs by the action of hemodynamic shear stresses on the endothelium. The transmission of hemodynamic forces throughout the endothelium and the mechanotransduction mechanisms that lead to biophysical, biochemical, and gene regulatory responses of endothelial cells to hemodynamic shear stresses are reviewed.
Figures
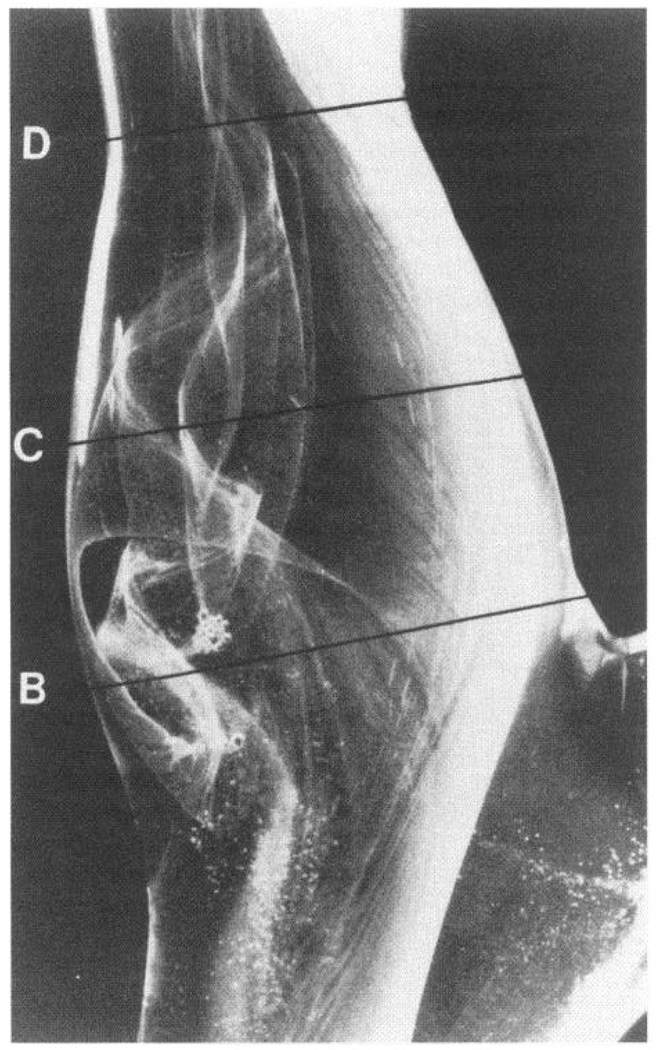
Glass model of human carotid sinus showing complex flow patterns visualized by hydrogen bubbles. In the sinus itself (B, C), there is separation of laminar flow profile to create a zone of recirculation that reattaches to main flow downstream (D). Vortices appear and disappear during pulsatile flow cycles. Note also helical pattern in main flow and absence of flow separation on opposite wall to sinus. Carotid artery atherosclerosis is common in region of flow separation. [From Zarins et al. (404).]

Outlines of endothelial cells in primate thoracic aorta adjacent to an intercostal branch artery. A region of predicted flow separation is shown in which shapes of cells change abruptly from elongated to polygonal (without preferred orientation). Polygonal morphology is believed to be caused by flow separation and vortices that subject endothelial surface to shear stress in several different directions during cardiac cycle. Adjacent cells are subjected to markedly different shear stress profiles. Gradients of shear stress are steepest on cells located at edge of flow separation region. Scanning electron micrograph is shown after silver staining.
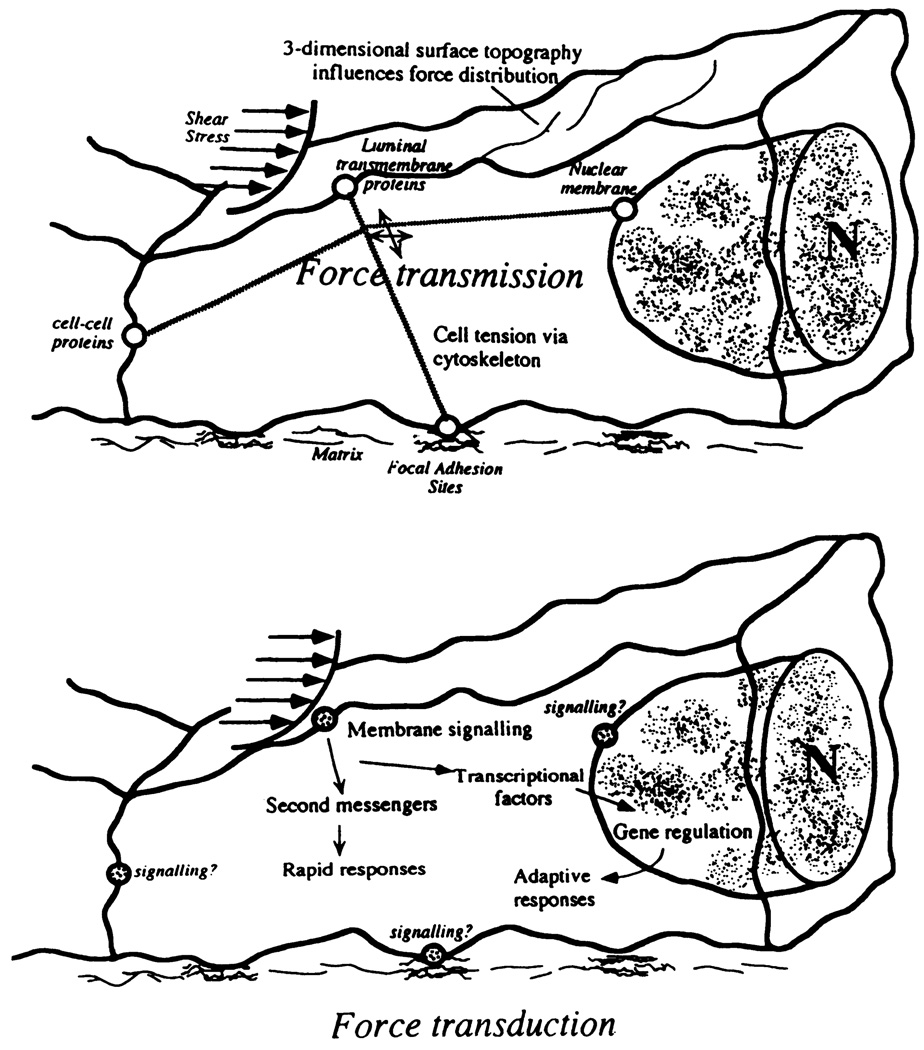
Separation of concepts of force transmission and force transduction. Cytoskeleton transfers stress to different regions of endothelial cell where mechanotransduction may occur. [From Davies and Barbee (61).]
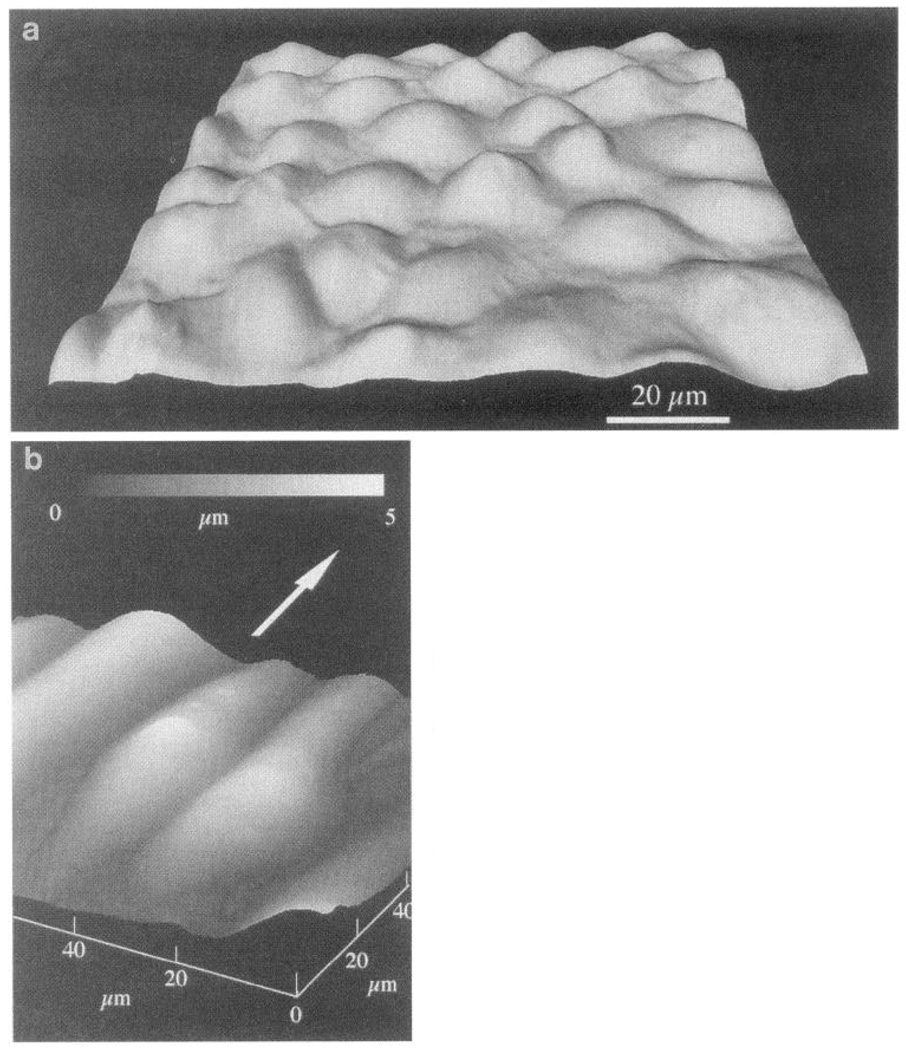
Atomic force microscope images of confluent bovine arotic endothelial cells. a: cultured under no-flow conditions. b: after 24-h exposure to 12-dyn/cm2 shear stress in steady laminar flow. Flattened profiles of aligned cells give rise to smaller spatial variations of shear stress and stress gradients at cell surface. Arrow indicates direction of flow. Gray micrometer scale in b indicates height of cell surface referenced to lowest point (at junctions between cells). (Images courtesy of Dr. Kenneth Barbee, University of Chicago, Chicago, IL.)

F-actin stress fiber distribution in aligned confluent endothelial cells. NBD-phallacidin stain was used. Arrow shows flow direction. [From Davies et al. (70). Reproduced from The Journal of Clinical Investigation, 1993, vol. 91, p. 2640–2652 by copyright permission of The Society for Clinical Investigation.]
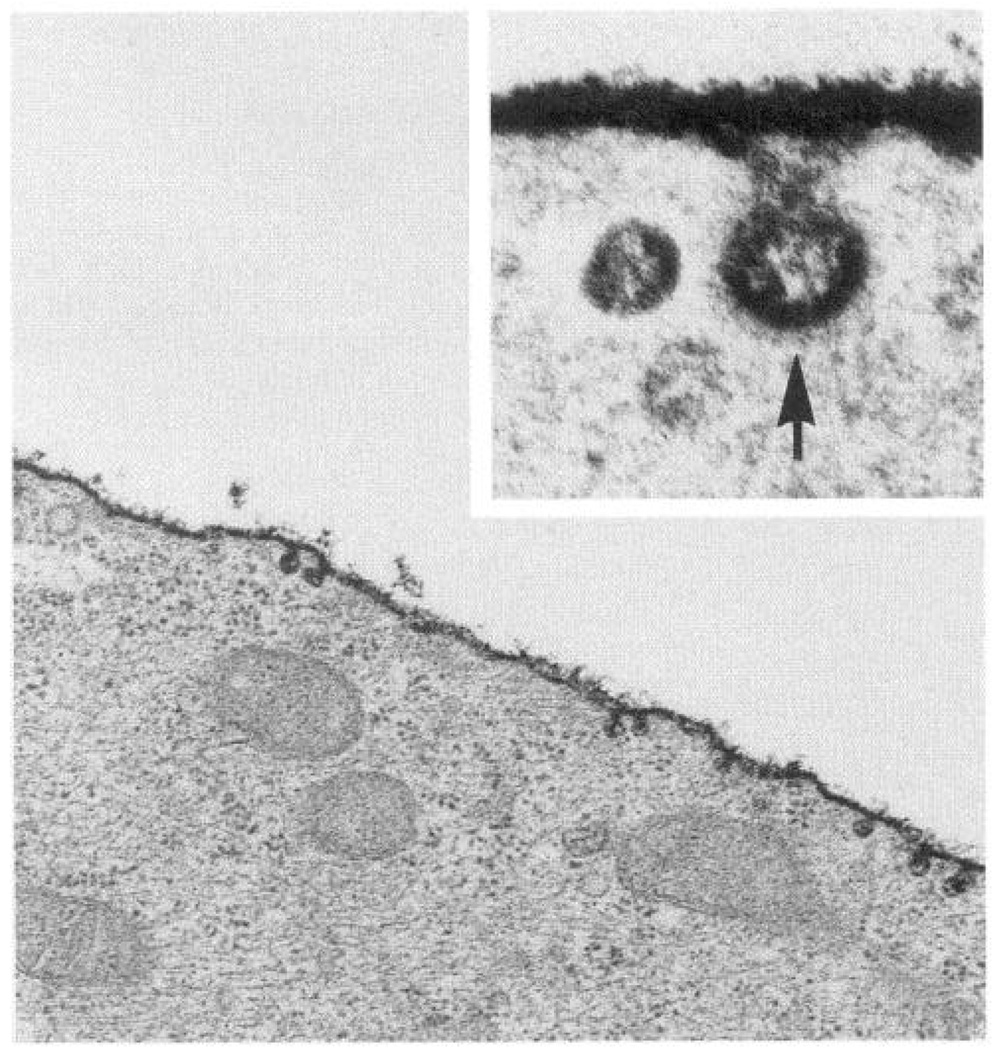
Endothelial cell luminal plasma membrane glycocalyx stained with ruthenium red. Inset: invagination of surface at a clathrin-coated pit. Transmission electron microscope magnification: ×40,000; inset, ×175,000.
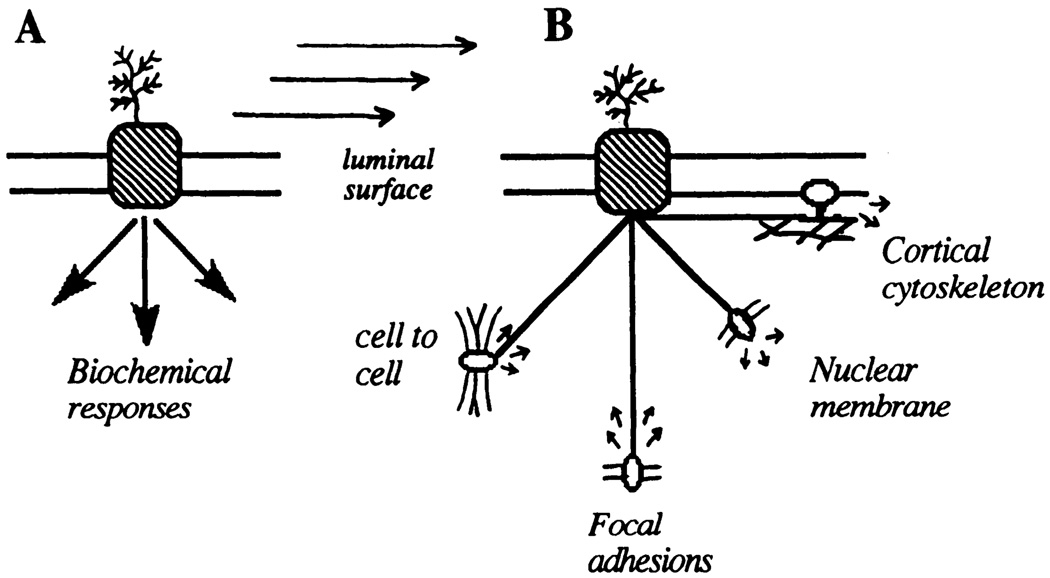
Centralized and decentralized mechanotransduction models in endothelial cells. A: shear stress transduction occurs at a primary site on luminal cell surface after which biochemical/electrophysiological pathways control responses through serial and parallel pathways. B: shear stress transduction occurs at multiple sites that are mechanically coupled by cytoskeleton. Each site, with different sensitivities, response times, and local environment, can generate bioresponses.
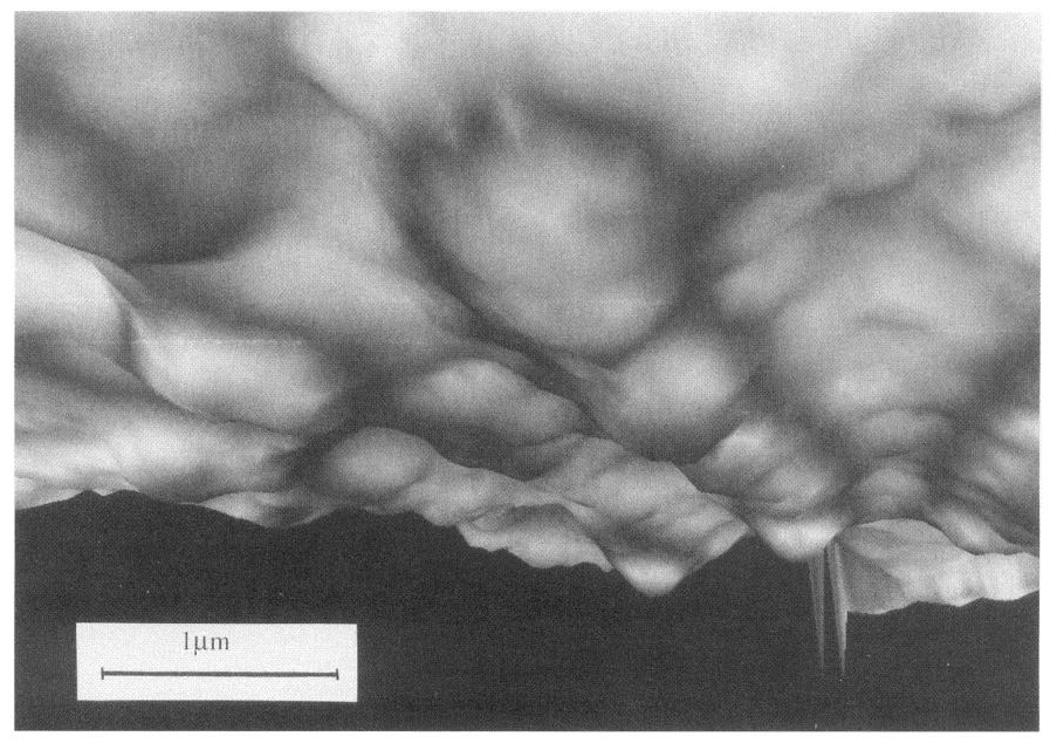
Endothelial abluminal cell surface geometry observed in real time by tandem scanning confocal microscopy after image processing (Quantex QX-7) and computer enhancement (Silicon Graphics Indigo II, IDL software). Membrane is organized into focal adhesion sites that extend downward to (invisible) substratum. Because this is a perspective view, scale marker refers to center of image only. (Courtesy of A. Robotewskyj and Dr. M. L. Griem, The University of Chicago, Chicago, IL.)
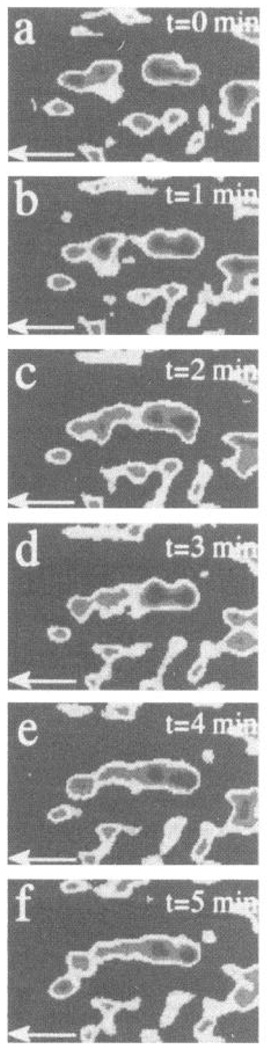
Tandem scanning confocal microscopy of endothelial cell focal adhesion site rearrangement during flow. Detailed real-time images 1 min apart of reorganizing focal contacts in a cell within a confluent monolayer during flow. Width of field is 2.5 µm. Progressive fusion and alignment of regions occurred over a short interval. Note that small area to left of newly forming aligned site did not significantly change, whereas other sites, above and below, migrated in direction of flow (shown by arrow). Cell boundaries did not significantly change during this period. [From Davies et al. (70). Reproduced from The Journal of Clinical Investigation, 1993, vol. 91, p. 2640–2652 by copyright permission of The Society for Clinical Investigation.]
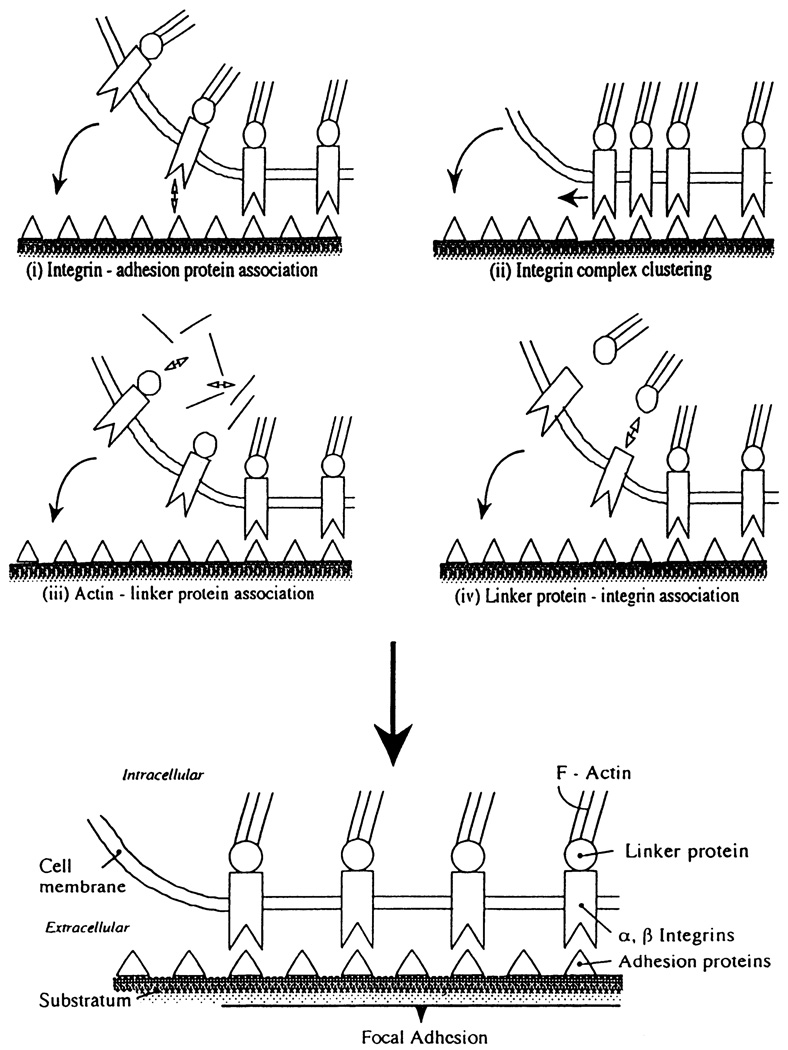
Mechanisms of focal adhesion formation/remodeling. Changes in affinities of principal proteins include those between integrins and adhesion proteins (i); between cytoskeleton and linker proteins such as talin, vinculin, tensin, paxillin, p125FAK, and α-actinin (iii); and between linker proteins and integrins (iv). Clustering of integrin complexes (ii) may also be involved.
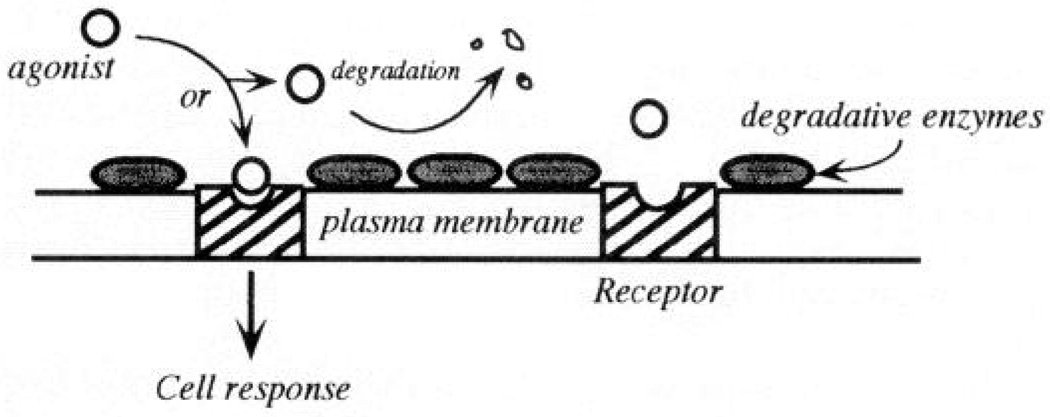
Mass transport at endothelial surface, Availability of an agonist in boundary layer adjacent to cell surface is determined by diffusion and convective delivery, balanced by removal via degradation and binding/internalization. If degradation is rapid, mass transport becomes rate limiting. Flow influences rate constants for mass transport and therefore indirectly regulates agonist availability to the cell.
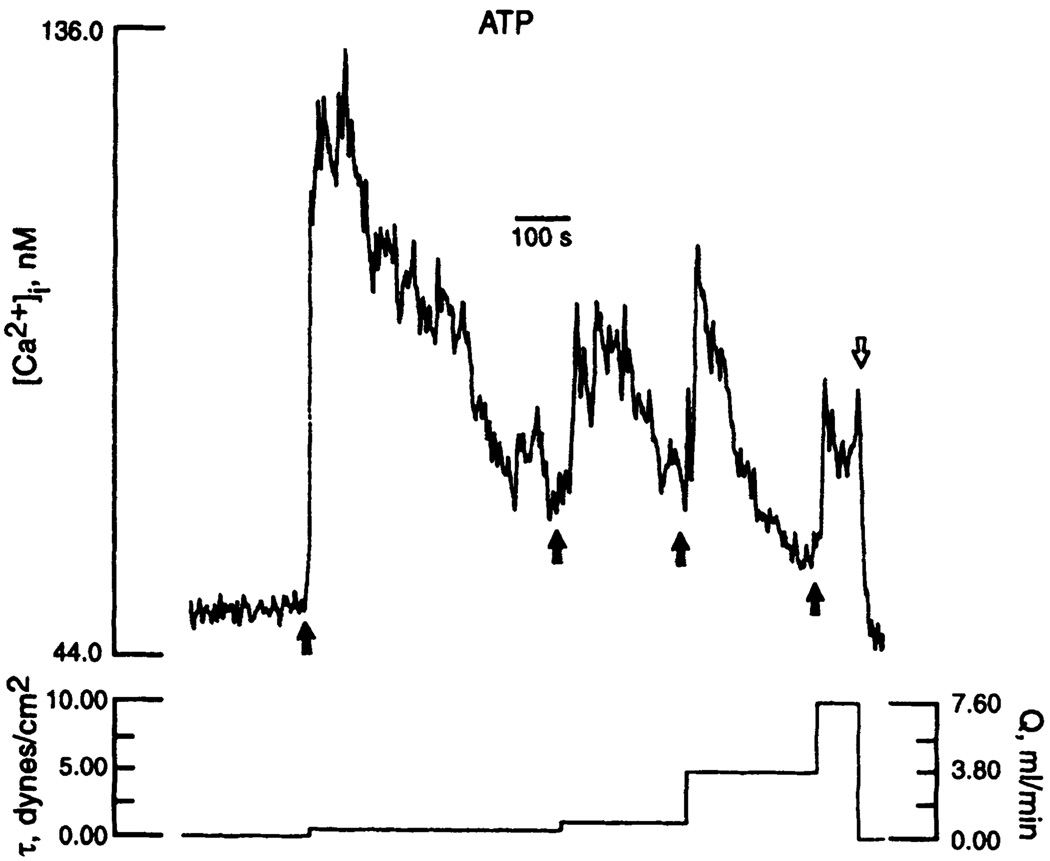
Flow modulates intracellular calcium ([Ca2+]i) response to purinergic agonist (extracellular ATP) in endothelial cells. A small change in flow rate (Q) [and shear stress (τ), dyn/cm2] disturbs mass transport of ATP in boundary layer, thereby favoring ATP-P2y purinoceptor interaction, which induces a calcium response. Concentration of ATP in bulk flow was constant at 1 mM. Up arrows indicate step increase of flow rate; down arrows indicate cessation of flow. [From Dull et al. (83).]
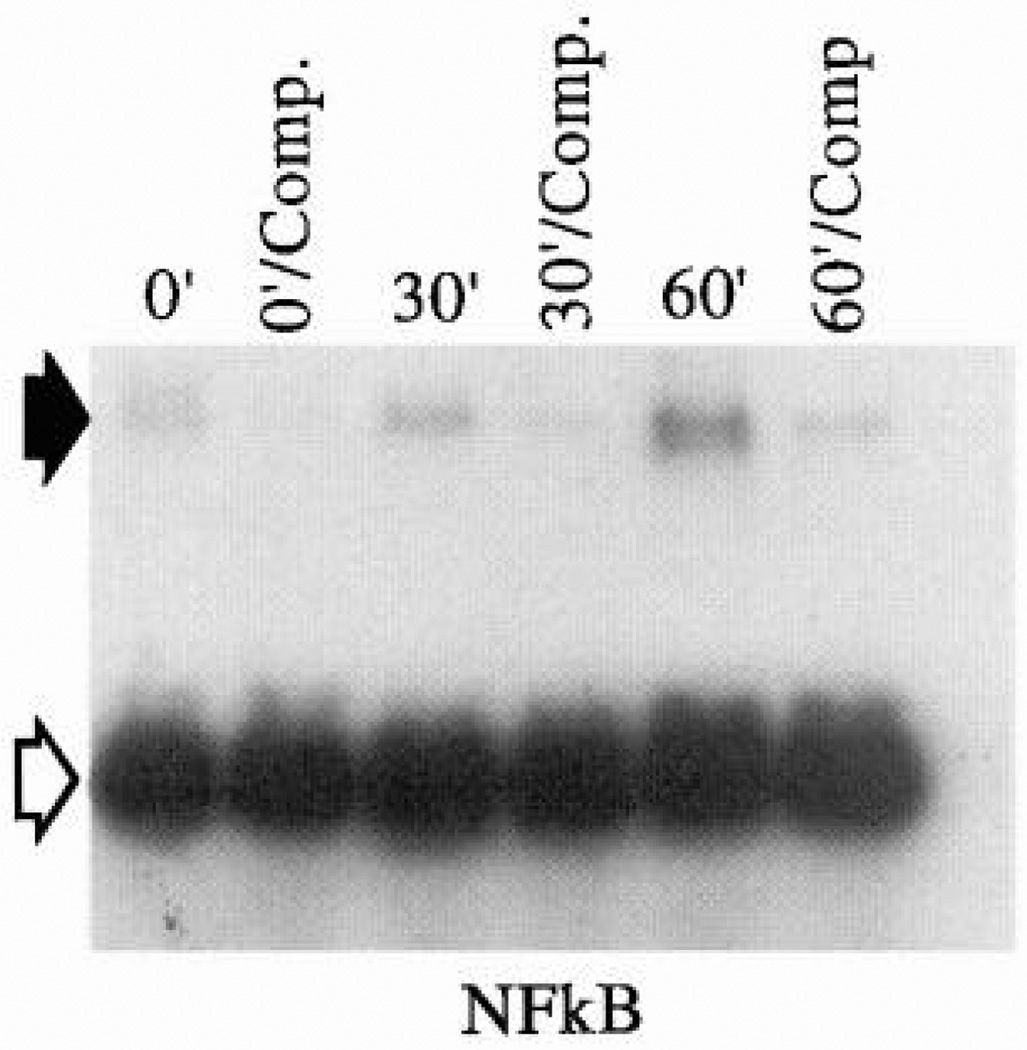
Nuclear factor κ B (NFκB)-DNA complex formation is stimulated by shear stress. Electrophoretic mobility shift assay of nuclear extracts isolated from confluent bovine aortic endothelial cells immediately after exposure to unidirectional laminar flow is shown. Control cells (lane 0') were placed in apparatus without flow. Other monolayers were exposed to steady shear stress of 12 dyn/cm2 for 30 and 60 min. Cell extracts were incubated with radiolabeled oligonucleotide containing consensus sequence for κB in the presence of nonspecific DNA-binding inhibitor poly(dI)-(dC), Extracts were incubated in absence (t') and presence (t'/comp) of a 100-fold molar excess of unlabeled oligonucleotide. DNA-protein complexes were resolved in 4% polyacrylamide gels, and gel shifts were documented by autoradiography. Closed arrow, NFκB-DNA complexes; open arrow, free κB oliognucleotide. [From Lan et al. (186).]
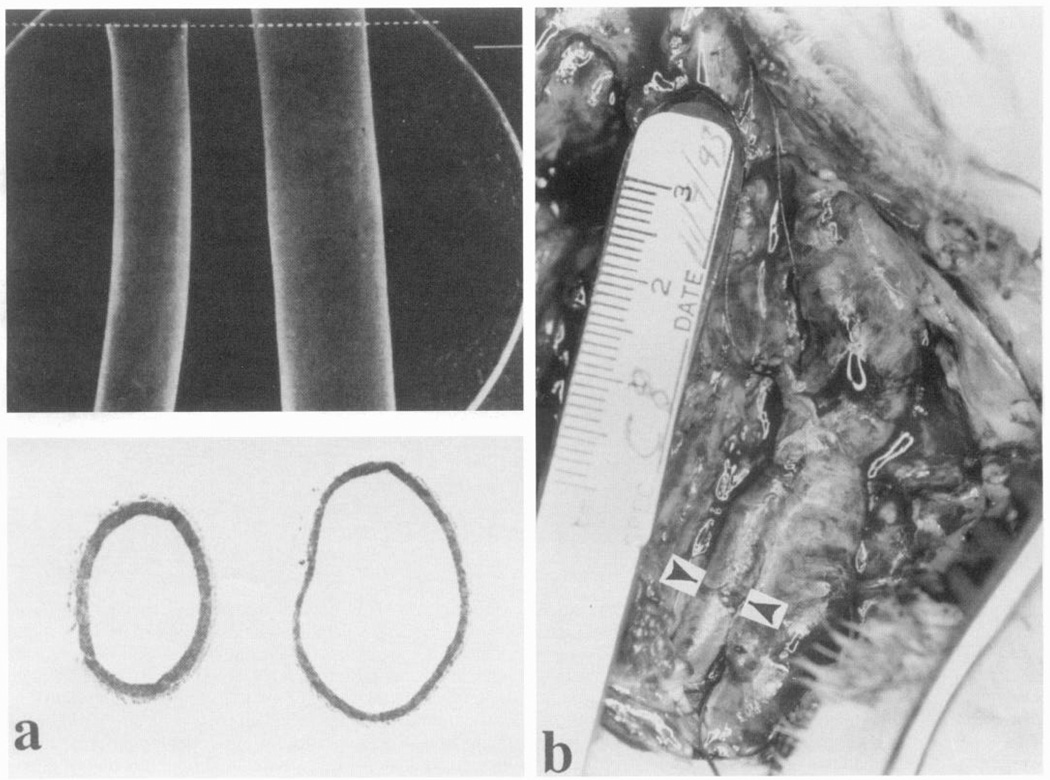
Arterial wall remodeling as a consequence of chronic alterations in flow. a: substantial decrease of rabbit carotid artery diameter 6 wk after flow had been reduced by coarctation. Top panel: vascular casts. Bottom panel: histological sections. Top and bottom panels show artery from treated and contralateral sides, respectively. Effects were dependent on presence of endothelium. [From Langille and O’Donnel (189).] b: increased diameter of rabbit carotid artery caused by an abnormally high flow rate by an arteriovenous fistula. Blood flow was increased 20-fold, resulting in a 4-fold increase in arterial diameter after 2 wk. Arrowheads indicate artery diameter. (Photograph courtesy of Dr. James McKinsey, Department of Surgery, University of Chicago, Chicago, IL.)
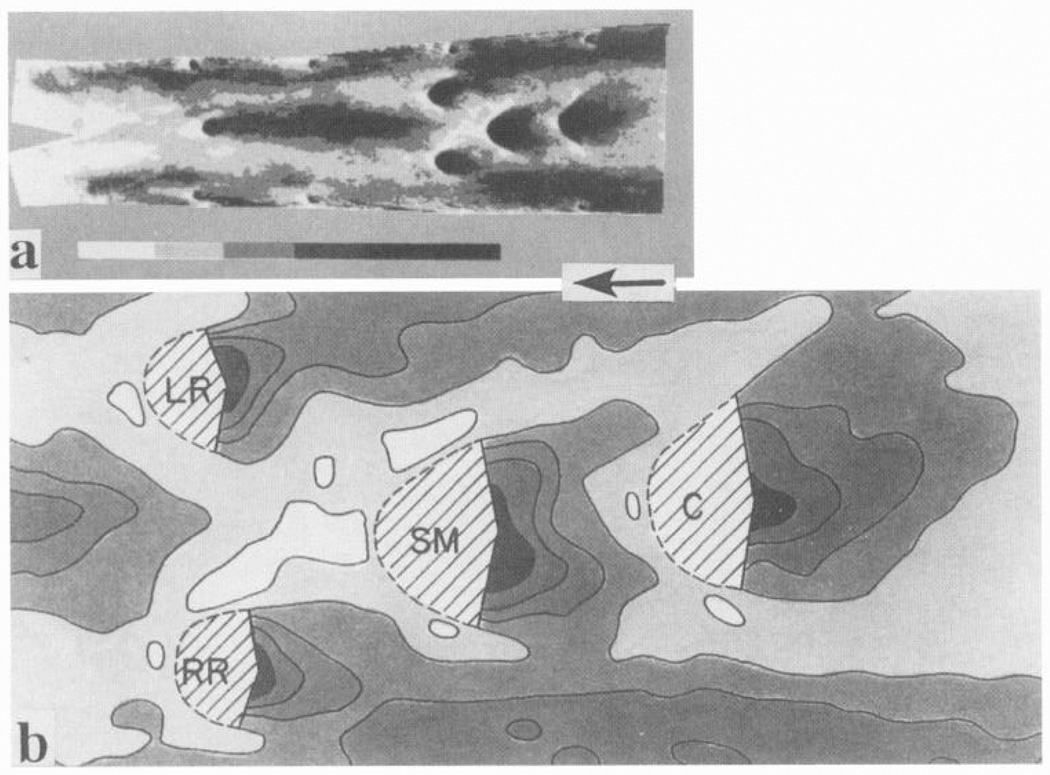
Localization of human atherosclerosis in abdominal aorta. a: digitization of sudanophilia intensity demonstrates lesions to be associated with regions of predicted complex hemodynamic profiles near branch arteries. b: detail of a showing distribution near right and left renal arteries (RR, LR) as well as superior mesenteric (SM) and celiac branches (C). Arrow indicates overall direction of flow, although detailed flow is complex. [From Cornhill et al. (52).]
References
-
- Achard C, Aynaud M. Recherches sur l’impregnation histologique de l’endothelium. Arch. Med. Exp. 1907;19:437–450.
-
- Adams DJ, Barakeh J, Laskey R, Van Breemen C. Ion channels and regulation of intracellular calcium in vascular endothelial cells. FASEB J. 1989;3:2389–2400. - PubMed
-
- Aderem A. The MARCKS brothers: a family of protein kinase C substrates. Cell. 1992;71:713–716. - PubMed
-
- Agard DA, Sedat JW. Three-dimensional architecture of a polytene nucleus. Nature Lond. 1983;302:676–681. - PubMed
-
- Ahmad M, Marui N, Alexander RW, Medford RM. Cell type specific transactivation of the VCAM-1 promoter through an NFκB enhancer motif. J. Biol. Chem. In press. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources