

Structures of the native and swollen forms of cowpea chlorotic mottle virus determined by X-ray crystallography and cryo-electron microscopy
- PMID: 7743132
- PMCID: PMC4191737
- DOI: 10.1016/s0969-2126(01)00135-6
Structures of the native and swollen forms of cowpea chlorotic mottle virus determined by X-ray crystallography and cryo-electron microscopy
Abstract
Background: RNA-protein interactions stabilize many viruses and also the nucleoprotein cores of enveloped animal viruses (e.g. retroviruses). The nucleoprotein particles are frequently pleomorphic and generally unstable due to the lack of strong protein-protein interactions in their capsids. Principles governing their structures are unknown because crystals of such nucleoprotein particles that diffract to high resolution have not previously been produced. Cowpea chlorotic mottle virions (CCMV) are typical of particles stabilized by RNA-protein interactions and it has been found that crystals that diffract beyond 4.5 A resolution are difficult to grow. However, we report here the purification of CCMV with an exceptionally mild procedure and the growth of crystals that diffract X-rays to 3.2 A resolution.
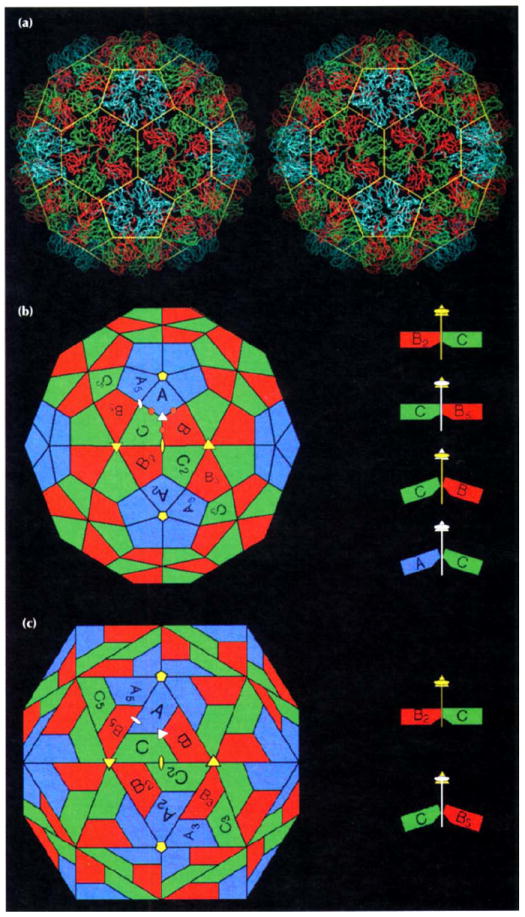
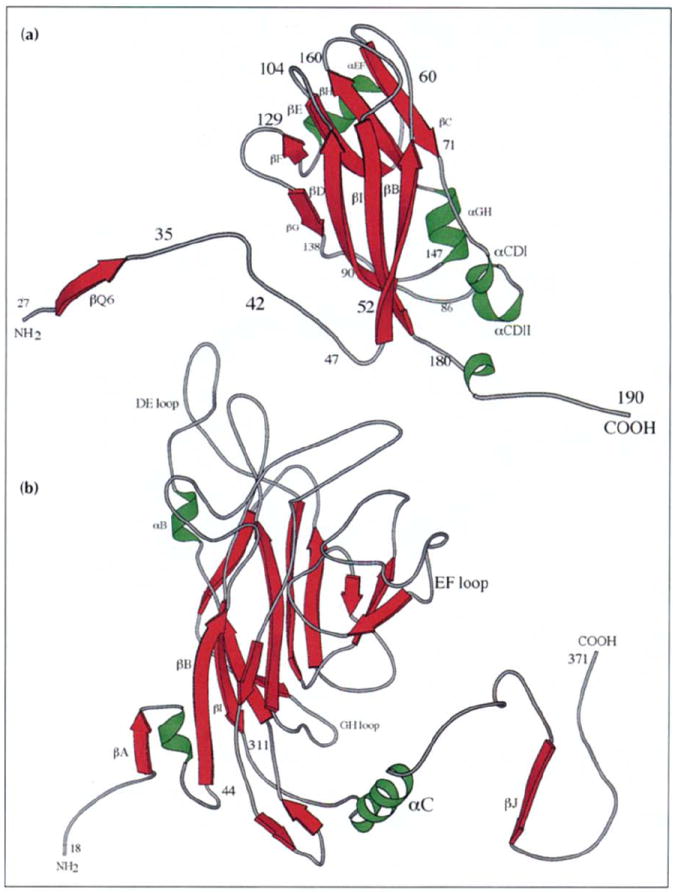
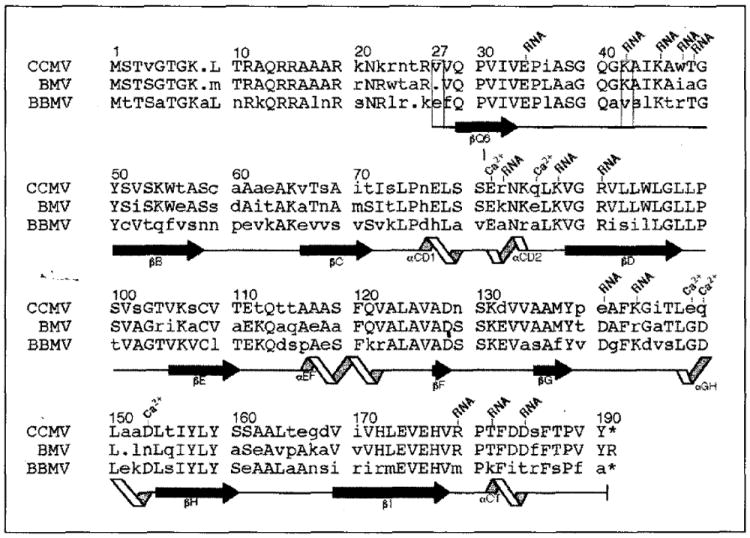
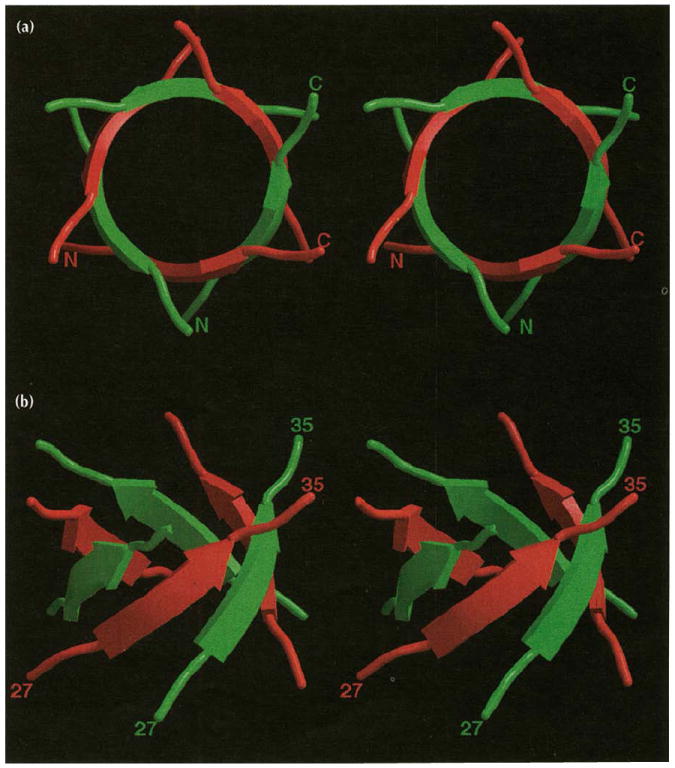
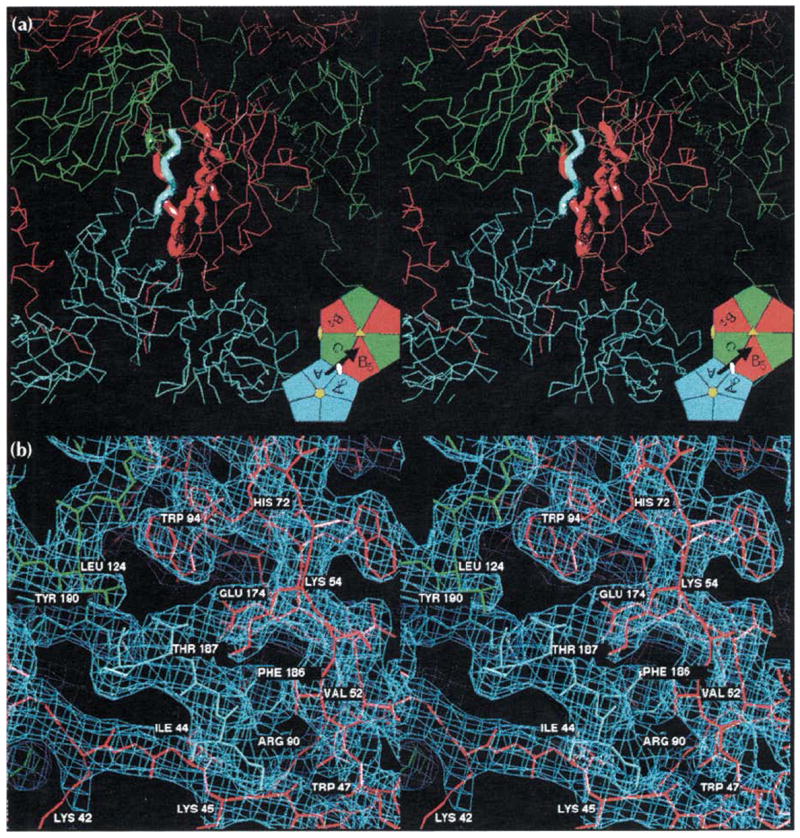
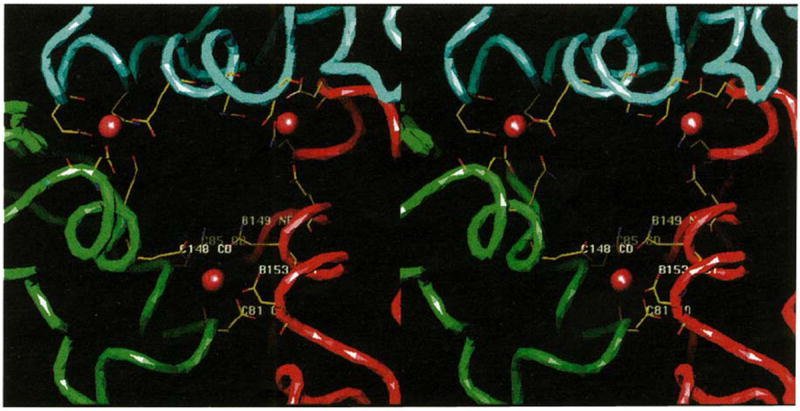
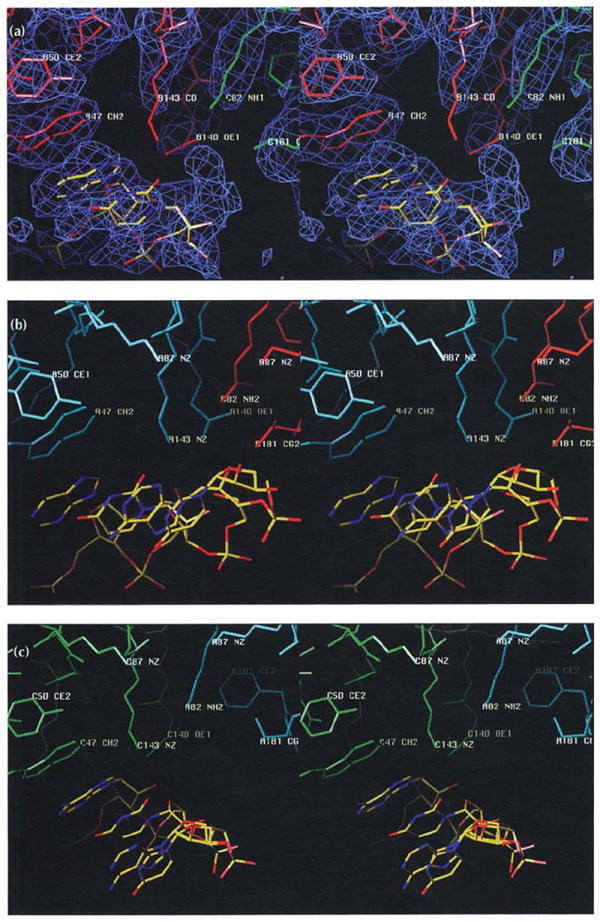
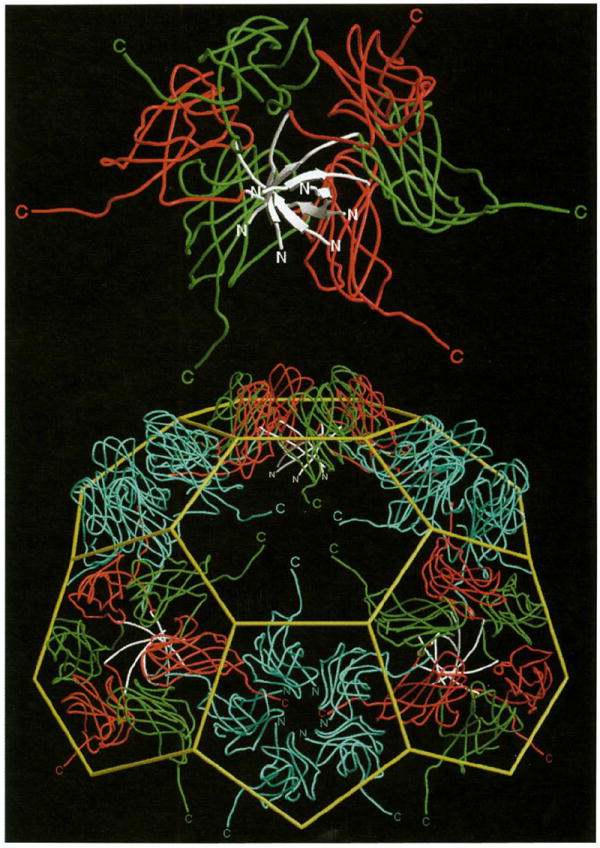
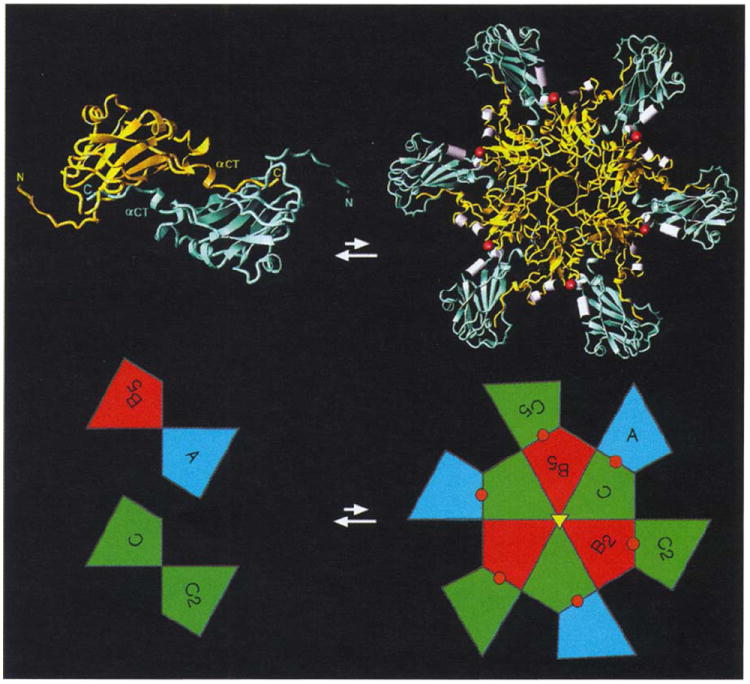
Results: The 3.2 A X-ray structure of native CCMV, an icosahedral (T = 3) RNA plant virus, shows novel quaternary structure interactions based on interwoven carboxyterminal polypeptides that extend from canonical capsid beta-barrel subunits. Additional particle stability is provided by intercapsomere contacts between metal ion mediated carboxyl cages and by protein interactions with regions of ordered RNA. The structure of a metal-free, swollen form of the virus was determined by cryo-electron microscopy and image reconstruction. Modeling of this structure with the X-ray coordinates of the native subunits shows that the 29 A radial expansion is due to electrostatic repulsion at the carboxyl cages and is stopped short of complete disassembly by preservation of interwoven carboxyl termini and protein-RNA contacts.
Conclusions: The CCMV capsid displays quaternary structural interactions that are unique compared with previously determined RNA virus structures. The loosely coupled hexamer and pentamer morphological units readily explain their versatile reassembly properties and the pH and metal ion dependent polymorphism observed in the virions. Association of capsomeres through inter-penetrating carboxy-terminal portions of the subunit polypeptides has been previously described only for the DNA tumor viruses, SV40 and polyoma.
Figures

References
-
- Speir A, Munshi S, Baker TS, Johnson JE. Preliminary X-ray data analysis of crystalline cowpea chlorotic mottle virus. Virology. 1993;193:234–241. - PubMed
-
- Francki RIB. The viruses and their taxonomy. In: Fraenkel-Conrat H, Wagner R, editors. The Plant Viruses. Plenum Press; New York: 1985. pp. 1–18.
-
- Lane LC. The bromoviruses. Adv Virus Res. 1974;19:151–220. - PubMed
-
- Baker TS. The packing of cowpea chlorotic mottle virus in crystalline monolayers. Proceedings of the Ninth International Congress on Electron Microscopy; 1978. pp. 24–25.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
