

Review
Nomenclature and classification of purinoceptors
Affiliations
- PMID: 7938164
- PMCID: PMC4976594
Item in Clipboard
Review
Nomenclature and classification of purinoceptors
Pharmacol Rev.
1994 Jun.
No abstract available
Figures

Amino acid sequences of purinoceptors deduced from cloned DNAs, aligned for maximum homology. Note that the P2 receptor sequences belong to a completely different family than do the adenosine (P1) receptors. The adenosine receptors listed here are all from the rat: A1 (Mahan et al., 1991), 326 amino acids; A2a (Fink et al., 1992; Furlong et al., 1992), 410 amino acids; A2b, (Stehle et al., 1992), 332 amino acids; A3 (Meyerhof et al., 1991; Zhou et al., 1992), 319 amino acids. Other such recombinant sequences known are for the human (Salvatore et al., 1992; Libert et al., 1992; Townsend-Nicholson and Shine, 1992), canine (Libert et al., 1991), and bovine (Olah et al., 1992; Tucker et al., 1992) A1 receptors; the human (Salvatore et al., 1992) and canine (Maenhaut et al., 1990) A2a receptors; and the human (Salvatore et al., 1992) A2b receptor, each of these is extremely homologous to the corresponding rat receptor. The rabbit A2 receptor was also found to be highly homologous, and genomic cloning revealed that the receptor gene has an intron (Bhattacharya et al., 1993). The chicken P2yl receptor has 362 amino acids (Webb et al., 1993), and the mouse P2u receptor (Lustig et al., 1993) has 373 amino acids. The approximate start positions of the transmembrane helices, as designated on the basis of hydropathy plots, are shown by the symbols TM1 to TM7.
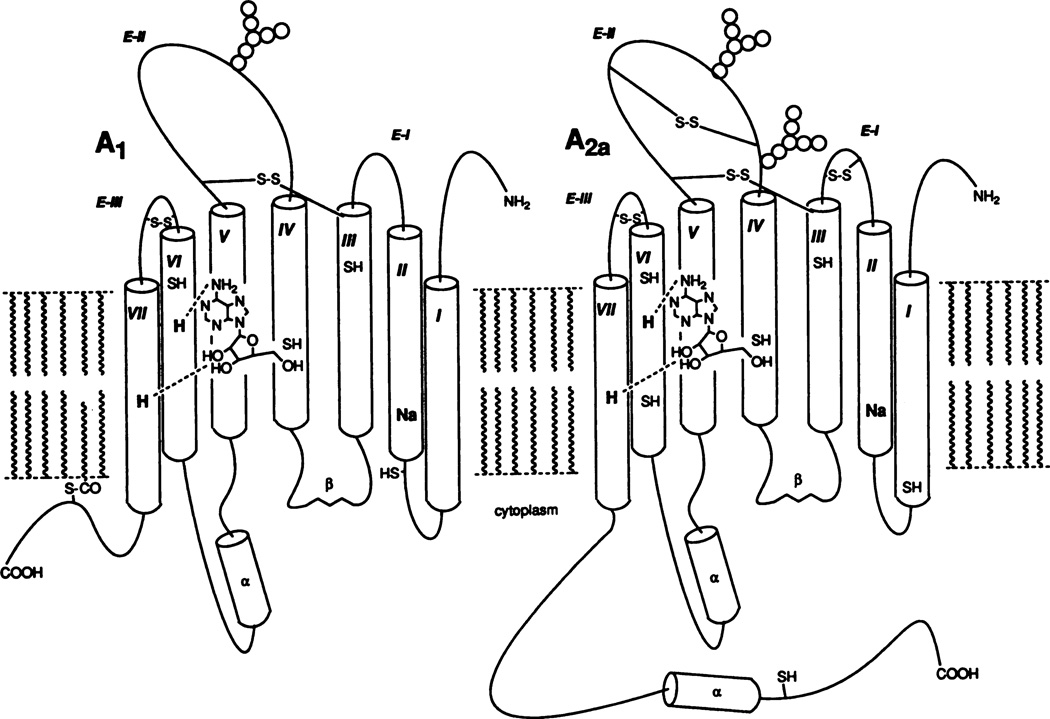
Proposed models of A1 and A2A adenosine receptor proteins based on sequence analysis (van Galen et al., 1992) and computer-assisted molecular modeling (Ijzerman et al., 1992). The seven transmembrane helices (I through VII) are arranged in a counterclockwise orientation (looking from the extracellular side) according to the experimentally determined structure of bacteriorhodopsin. Actually, I and VII are in proximity, forming a barrel shape, which surrounds the ligand-binding site. Histidyl residues (H) in the sixth and seventh helices are proposed to hydrogen bond to adenosine, through the purine N6- and ribose 2′,3′ positions, respectively. The locations of cysteinyl residues (SH) and hypothetical disulfide bridges (S-S, Jacobson et al., 1993d) are indicated. Glycosylation occurs on the second extracellular loop (E-II) in both receptors. In the A1 receptor, a potential palmitoylation site (S-CO) is shown as forming an additional anchor of the carboxy-terminal segment in the phospholipid bilayer. Cytoplasmic segments show a hypothetical secondary structure (α and β), predicted using computational algorithms (van Galen et al., 1992)
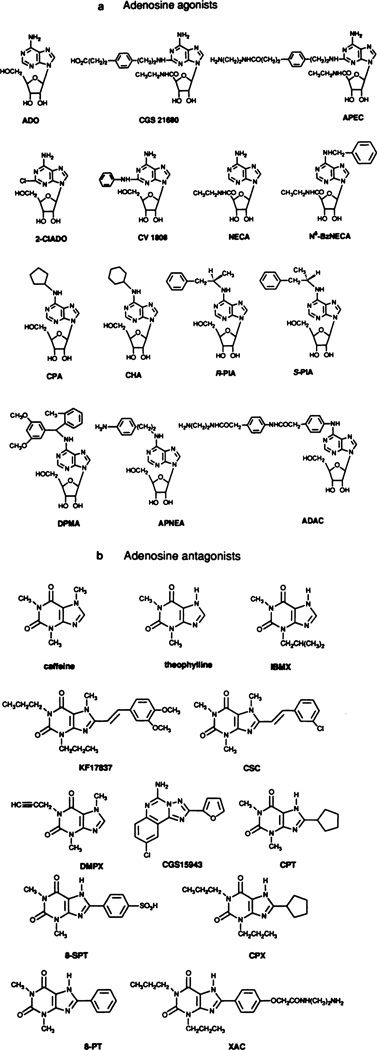
Ligands used to classify adenosine receptors. Adenosine agonists (a) include those that are selective for A1 receptors [N6-cyclopentyladenosine (CPA) > N6-cyclohexyladenosine (CHA) > N6(R-phenylisopropyl)-adenosine (R-PIA)], A2a receptors (2-[p-(2-car-bonylethyl)-phenylethylamino]-5′N-ethylcarboxamidoadenosine (CG21680)>2-[(2-aminoethylamino)carbonylethylphenylethylamino]-5′N-ethylcarboxamidoadenosine (APEC) > 2-phenylaminoadenosine (CV1808)), and A3 receptors [N6-benzyl-5′-N-ethyl-carboxamidoadenosine (NECA)]. NECA and 2-chloroadenosine (2-C1ADO) are essentially nonselective. Adenosine antagonists (b) include such that are A1 selective {l,3-dipropyl-8-cyclopentylxanthine (CPX, or sometimes DPCPX); 8-cyclopentyltheophylline (CPT) [and xanthine amine congener (XAC)] |, thosethatareA2a,selective[8-(3-chlorostryl)caffeine(CSC);1,3-dipropyl-8-(3,4-dimethoxystyryl)-7-methylxanthine (KF 17837)], moderately A2 selective [1,3-dimethyl-7-propylxanthine (DMPX) and 9-chloro-2-(2-furanyl)-5,6-dihydro-[1,2,4]-triazolo[1,5]quinazolin-5-imine monome-thanesulfonate(CGS 15943)}. Caffeine, theophylline, 3-isobutyl-1-methylxanthine (IBMX), and 8p-sulfophenyl theophylline are essentially nonselective. The latter compound penetrates the blood-brain barrier poorly. (For other abbreviations, see footnote to table 3).
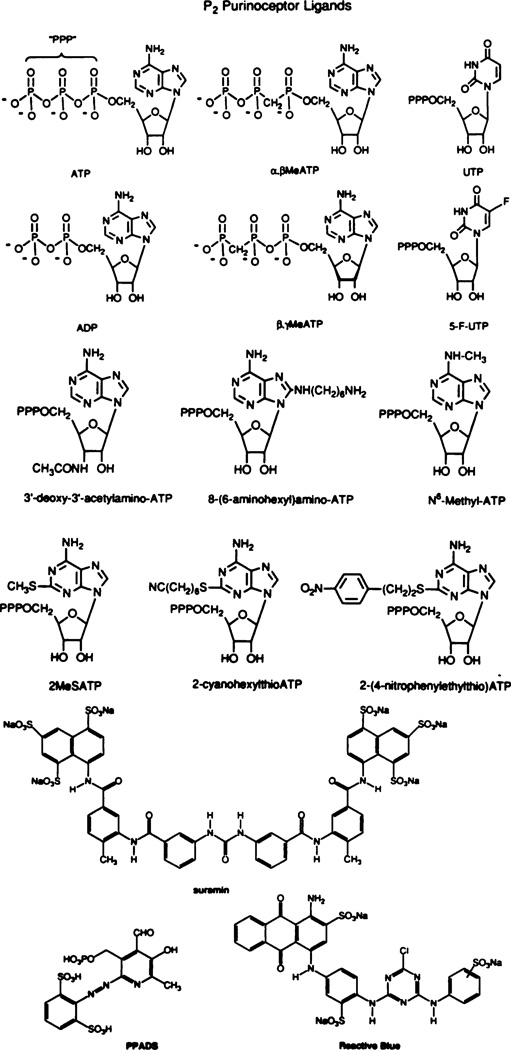
Ligands used to characterize P2 purinoceptors. The structures shown are of the d -isomers. Some studies have also been done on l -isomers. α,β-methylene (Me) ATP is a potent and selective P2X agonist. 8-(6-aminohexylamino)ATP is selective for P2Y vs. P2X receptors and may discriminate between subforms (Burnstock et al., 1994). UTP and 5-F-UTP are active at P2U but not at P2Y receptors. 2-MethylthioATP is active at P2Y but not at P2U receptors. None of these are very active at P2X receptors. The long-chain functionalized congeners (Fischer et al., 1993) 2-(p-nitrophenylthio)ATP and 2-(6-cyanhexylthio)ATP maintain or increase potency at P2Yreceptors. The nitro derivative may discriminate between forms of P2X receptors. N6-methyl ATP may discriminate between forms of P2Y receptors. The structures of some compounds—suramin, pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid (PPADS), and Reactive Blue—that have antagonistic properties are also shown.
References
-
- Abbracchio M, Burnstock G. Purinoceptors: are there families of P2X and P2Y purinoceptors? Pharmacol. Ther. in press. - PubMed
-
- Barnard EA, Burnstock G, Webb TE. G-protein coupled receptors for ATP and other nucleotides: a new receptor family. Trends Pharmacol. Sci. 1994;15:67–70. - PubMed
-
- Bean BP. Pharmacology and electrophysiology of ATP-activated ion channels. Trends Pharmacol. Sci. 1992;13:87–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
