

Direct observation of single kinesin molecules moving along microtubules
- PMID: 8602245
- PMCID: PMC2852185
- DOI: 10.1038/380451a0
Direct observation of single kinesin molecules moving along microtubules
Abstract
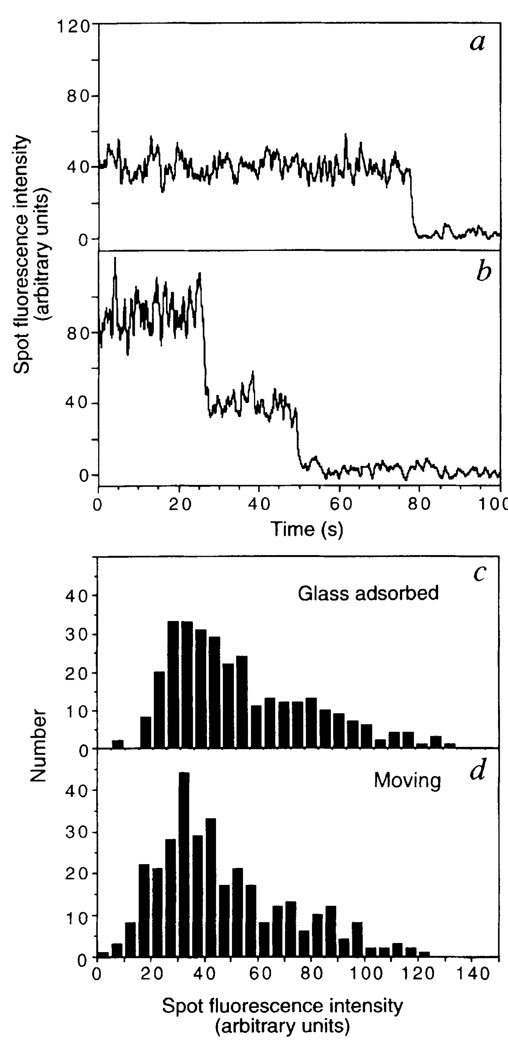
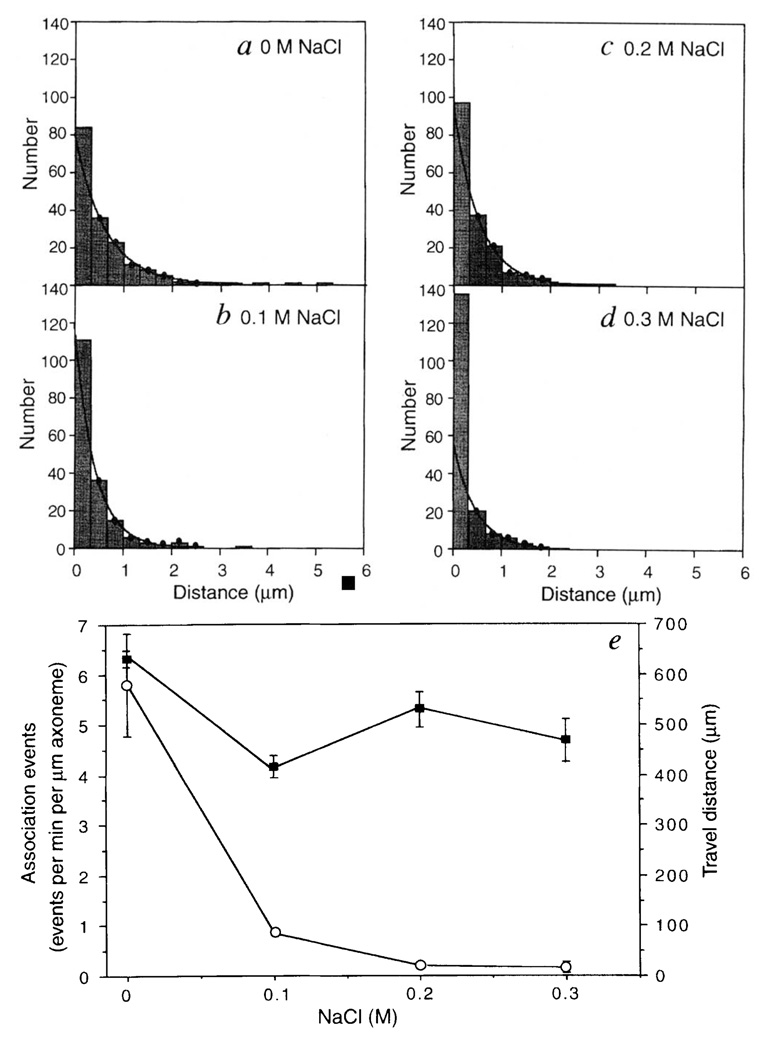
Kinesin is a two-headed motor protein that powers organelle transport along microtubules. Many ATP molecules are hydrolysed by kinesin for each diffusional encounter with the microtubule. Here we report the development of a new assay in which the processive movement of individual fluorescently labelled kinesin molecules along a microtubule can be visualized directly; this observation is achieved by low-background total internal reflection fluorescence microscopy in the absence of attachment of the motor to a cargo (for example, an organelle or bead). The average distance travelled after a binding encounter with a microtubule is 600 nm, which reflects a approximately 1% probability of detachment per mechanical cycle. Surprisingly, processive movement could still be observed at salt concentrations as high as 0.3 M NaCl. Truncated kinesin molecules having only a single motor domain do not show detectable processive movement, which is consistent with a model in which kinesin's two force-generating heads operate by a hand-over-hand mechanism.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
