

Layer-specific properties of the transient K current (IA) in piriform cortex
- PMID: 8656280
- PMCID: PMC6578623
- DOI: 10.1523/JNEUROSCI.16-12-03862.1996
Layer-specific properties of the transient K current (IA) in piriform cortex
Abstract
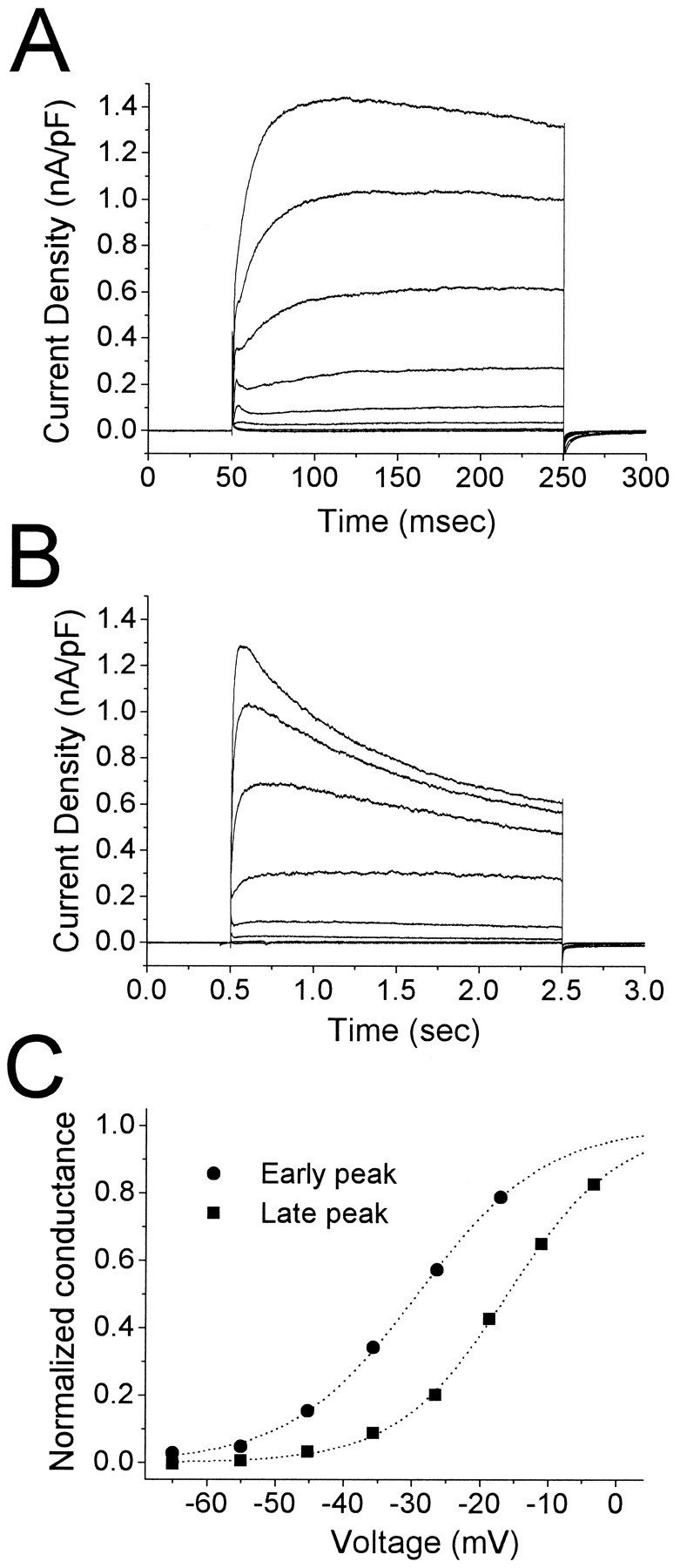
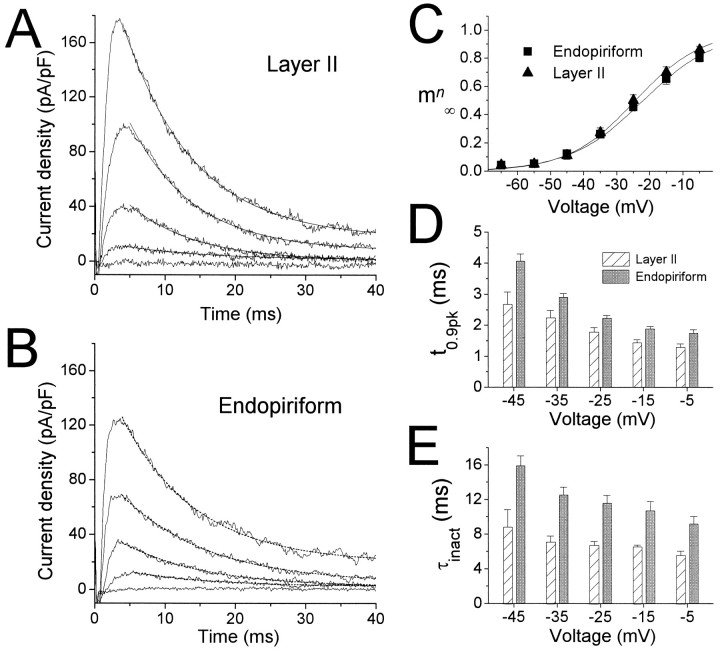
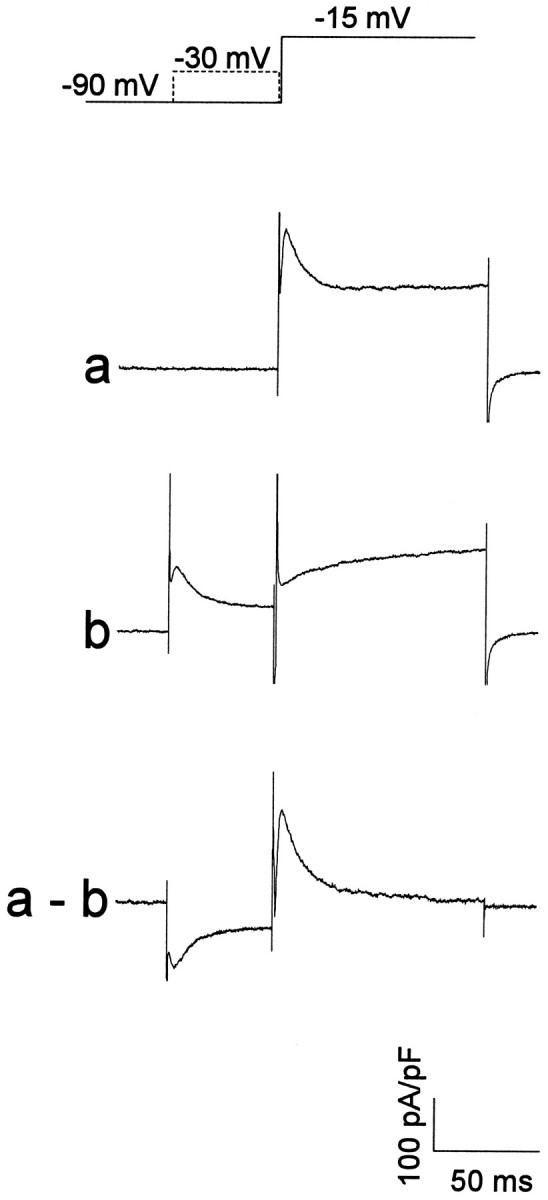
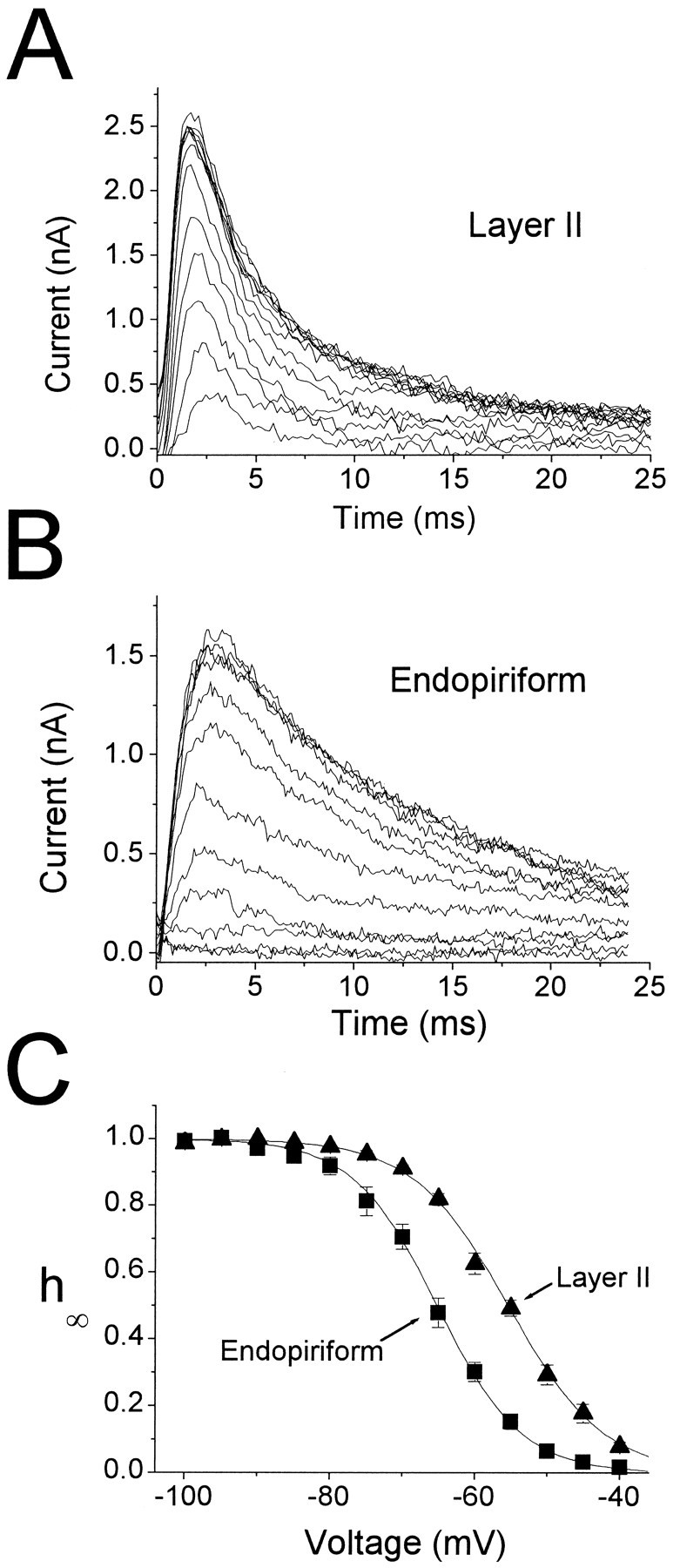
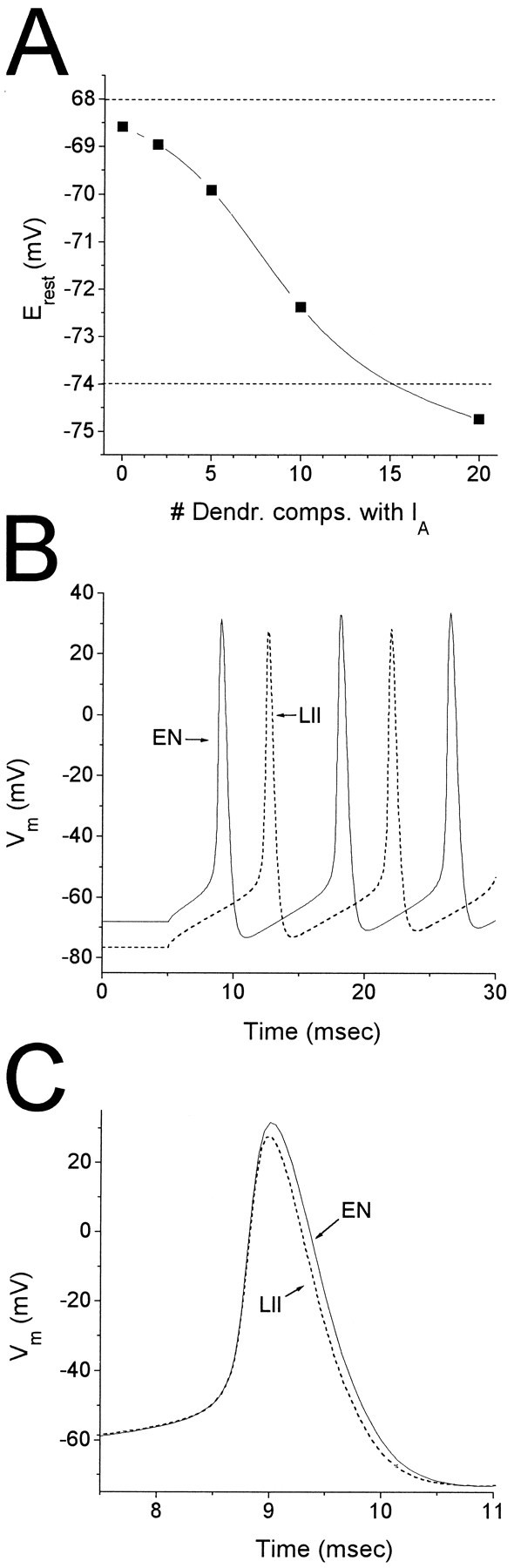
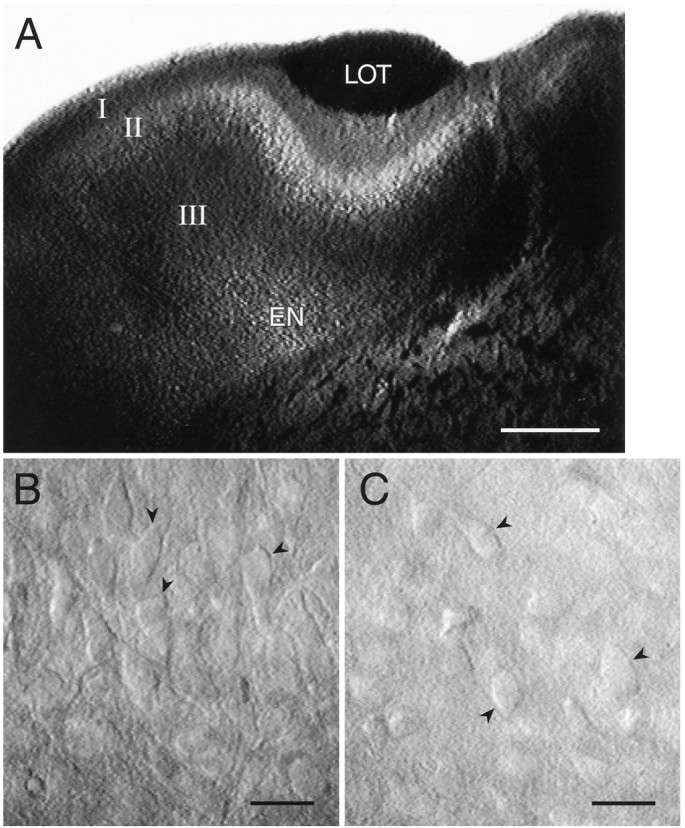
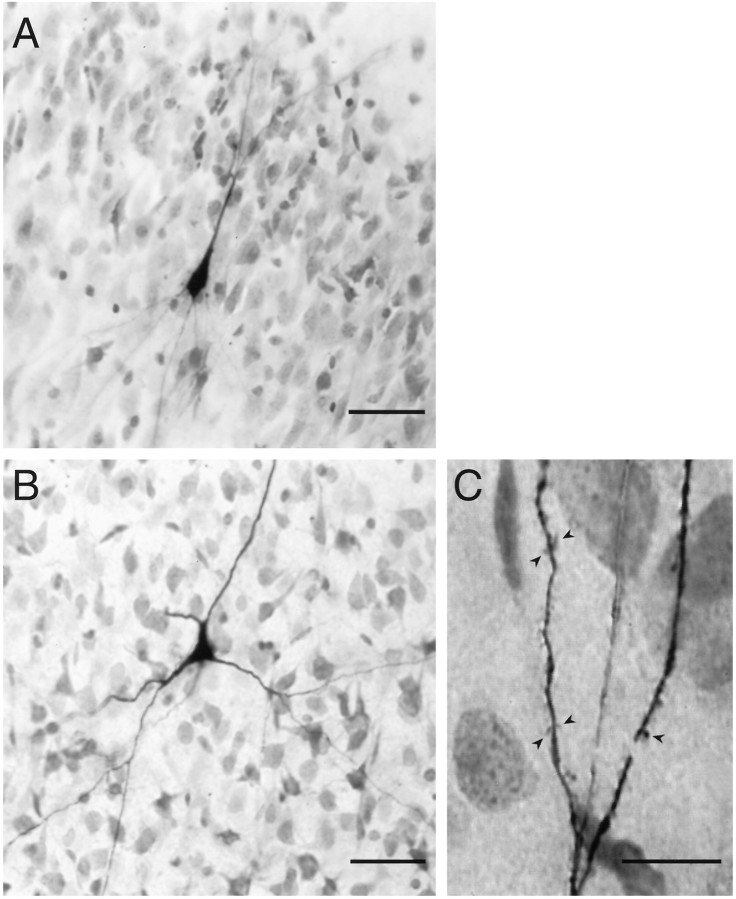
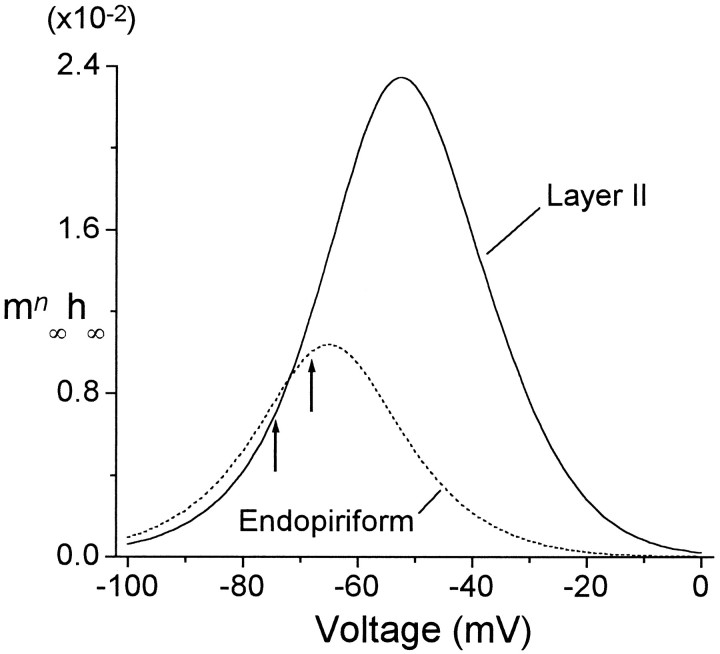
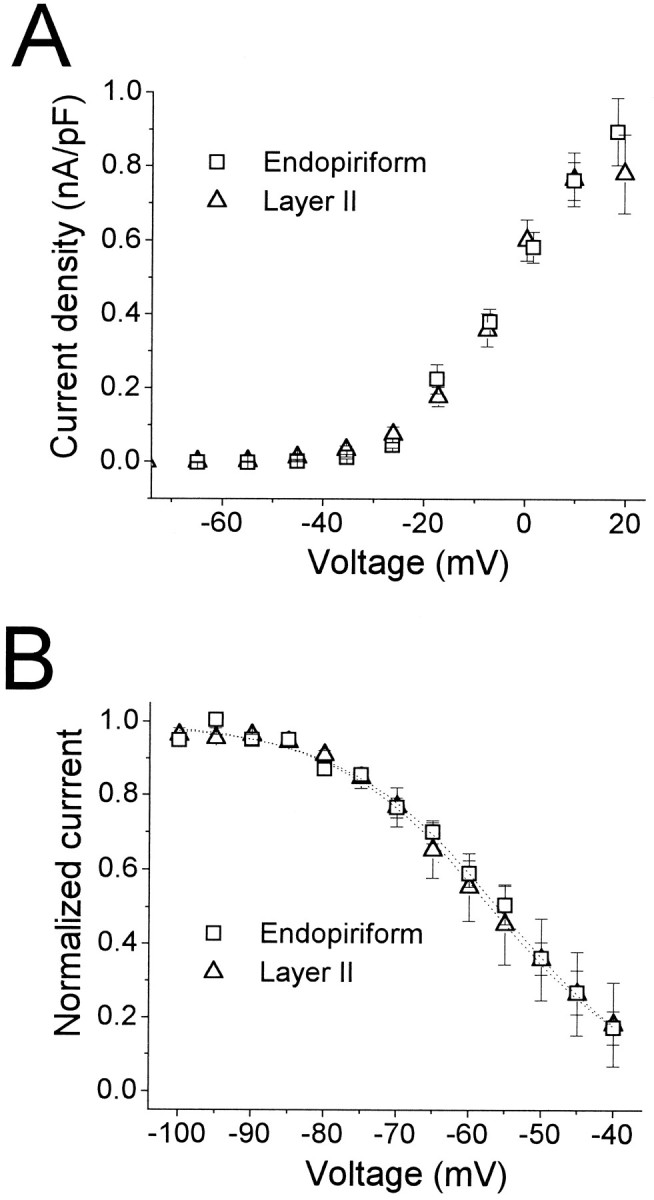
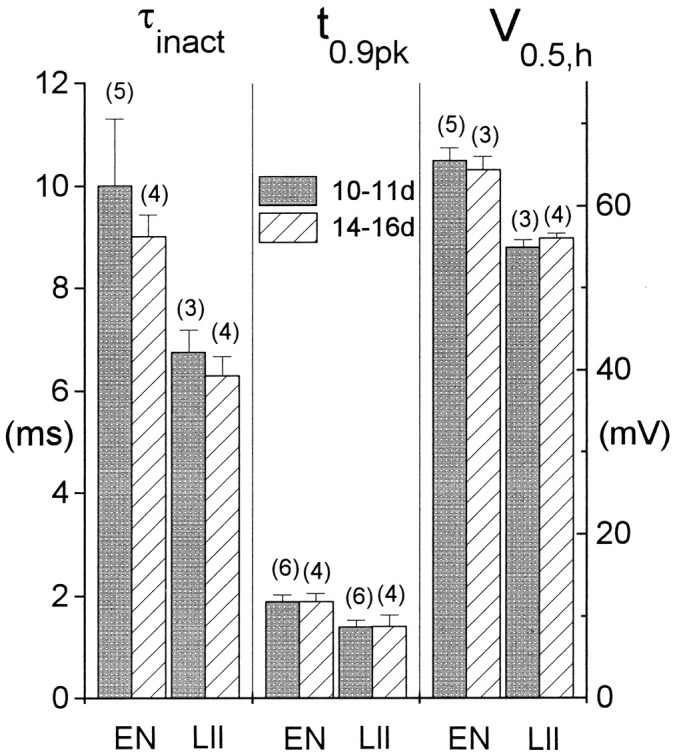
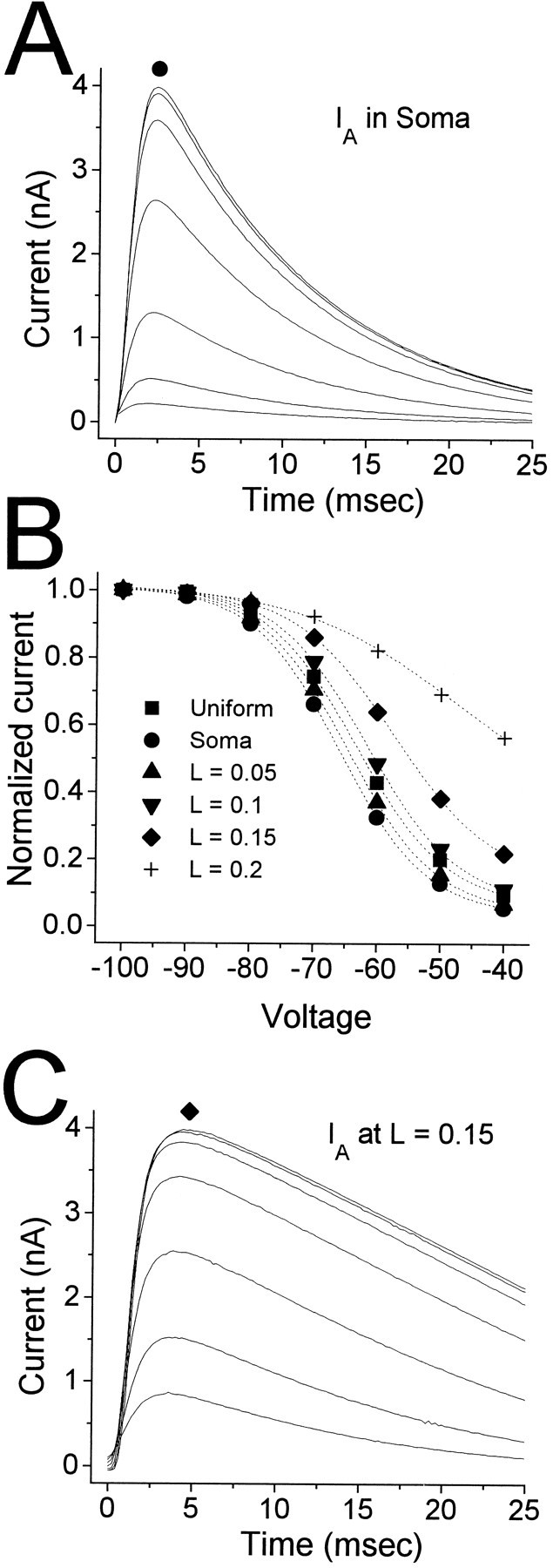
Piriform cortex in the rat is highly susceptible to induction of epileptiform activity. Experiments in vivo and in vitro indicate that this activity originates in endopiriform nucleus (EN). In slices, EN neurons are more excitable than layer II (LII) pyramidal cells, with more positive resting potentials and lower spike thresholds. We investigated potassium currents in EN and LII to evaluate their contribution to these differences in excitability. Whole-cell currents were recorded from identified cells in brain slices. A rapidly inactivating outward current (IA) had distinct properties in LII (IA,LII) versus EN (IA,EN). The peak amplitude of IA,EN was 45% smaller than IA,LII, and the kinetics of activation and inactivation was significantly slower for IA,EN. The midpoint of steady-state inactivation was hyperpolarized by 10 mV for IA,EN versus IA,LII, whereas activation was similar in the two cell groups. Other voltage-dependent potassium currents were indistinguishable between EN and LII. Simulations using a compartmental model of LII cells argue that different cellular distributions of IA channels in EN versus LII cells cannot account for these differences. Thus, at least some of the differences are intrinsic to the channels themselves. Current-clamp simulations suggest that the differences between IA,LII and IA,EN can account for the observed difference in resting potentials between the two cell groups. Simulations show that this difference in resting potential leads to longer first spike latencies in response to depolarizing stimuli. Thus, these differences in the properties of IA could make EN more susceptible to induction and expression of epileptiform activity.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials