

Organization of AMPA receptor subunits at a glutamate synapse: a quantitative immunogold analysis of hair cell synapses in the rat organ of Corti
- PMID: 8699256
- PMCID: PMC6578857
- DOI: 10.1523/JNEUROSCI.16-14-04457.1996
Organization of AMPA receptor subunits at a glutamate synapse: a quantitative immunogold analysis of hair cell synapses in the rat organ of Corti
Abstract
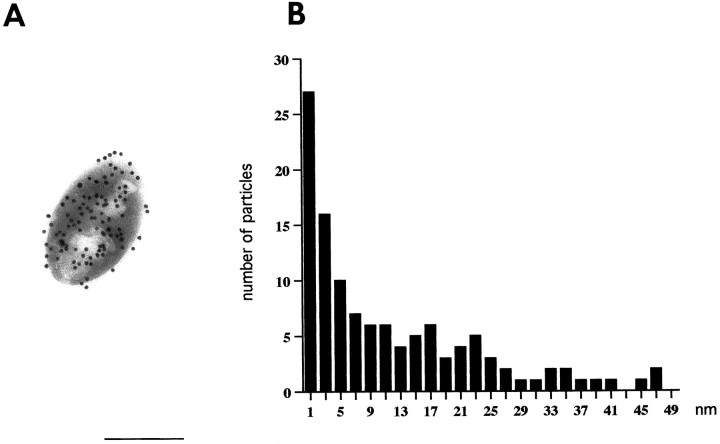
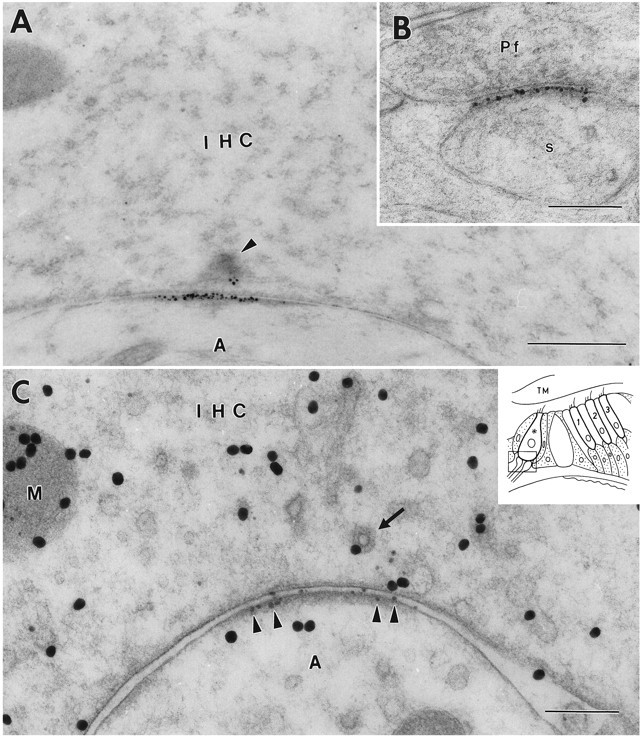
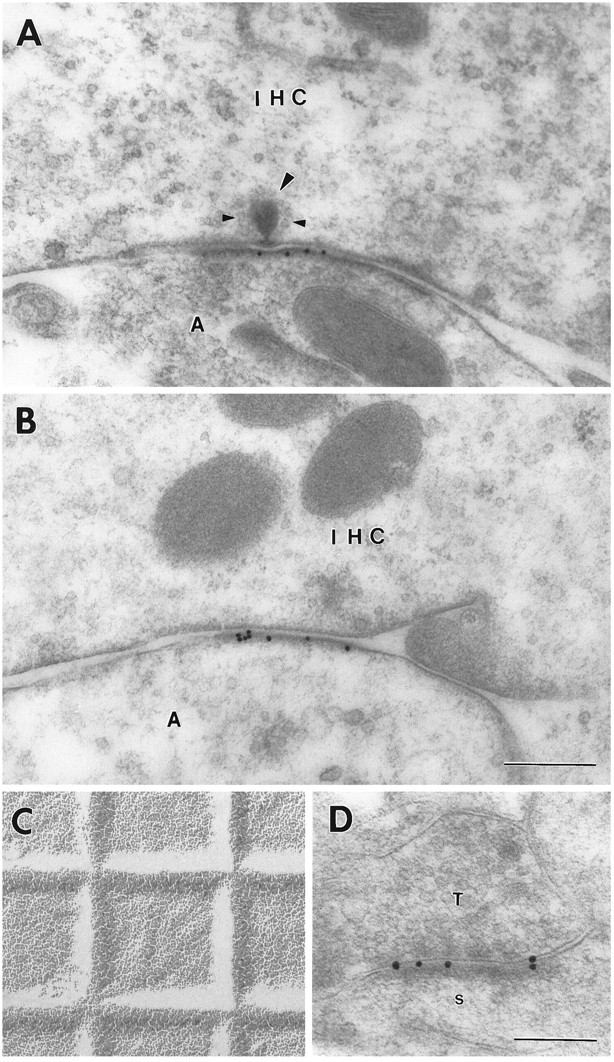
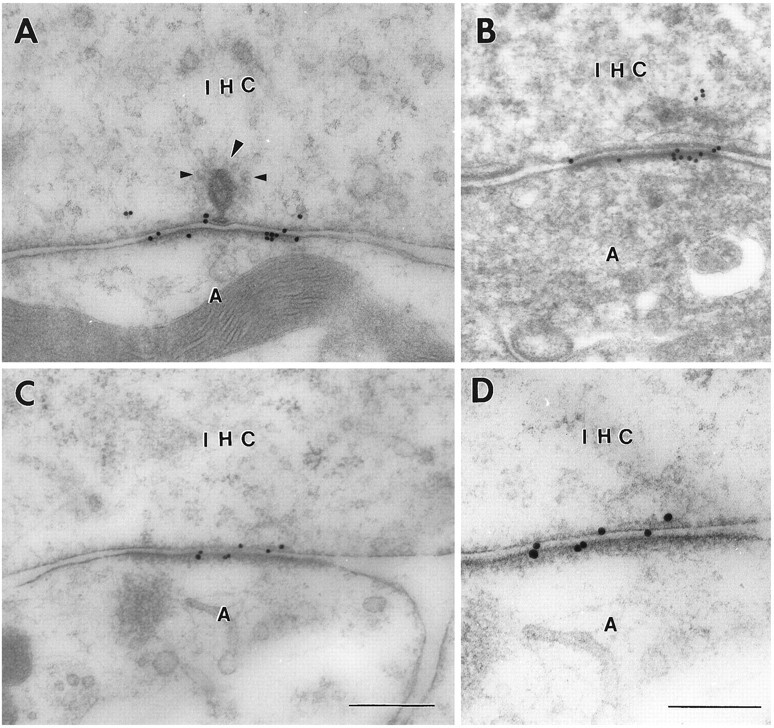
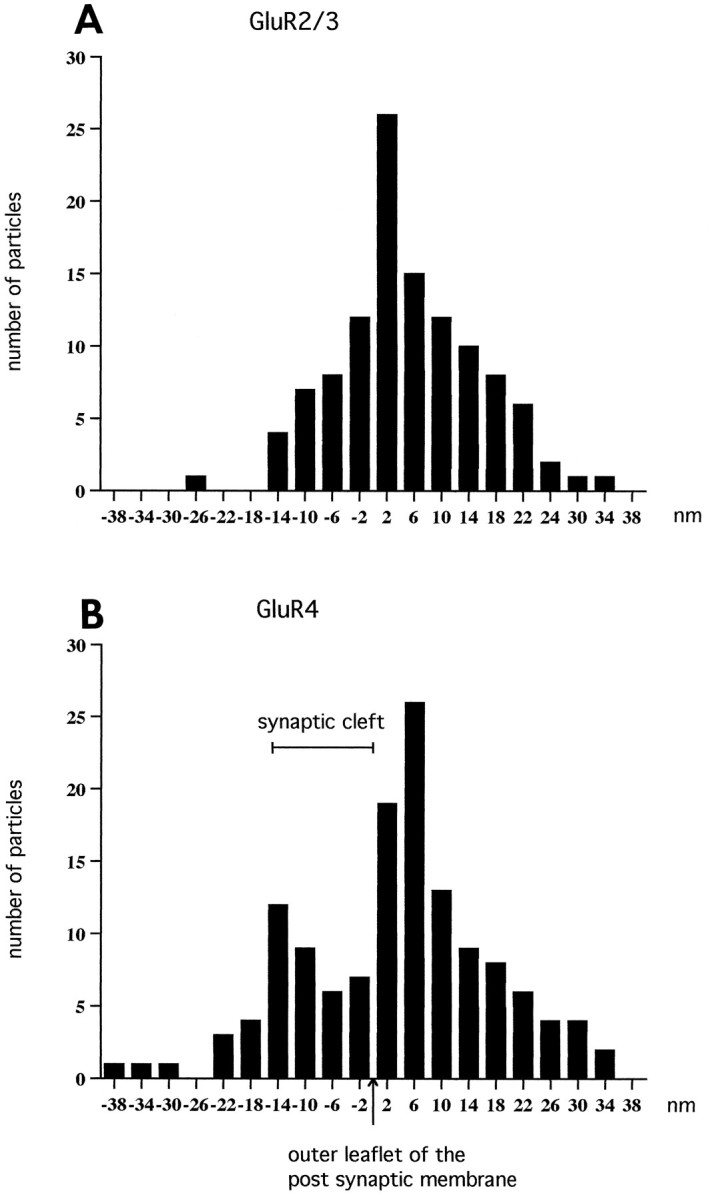
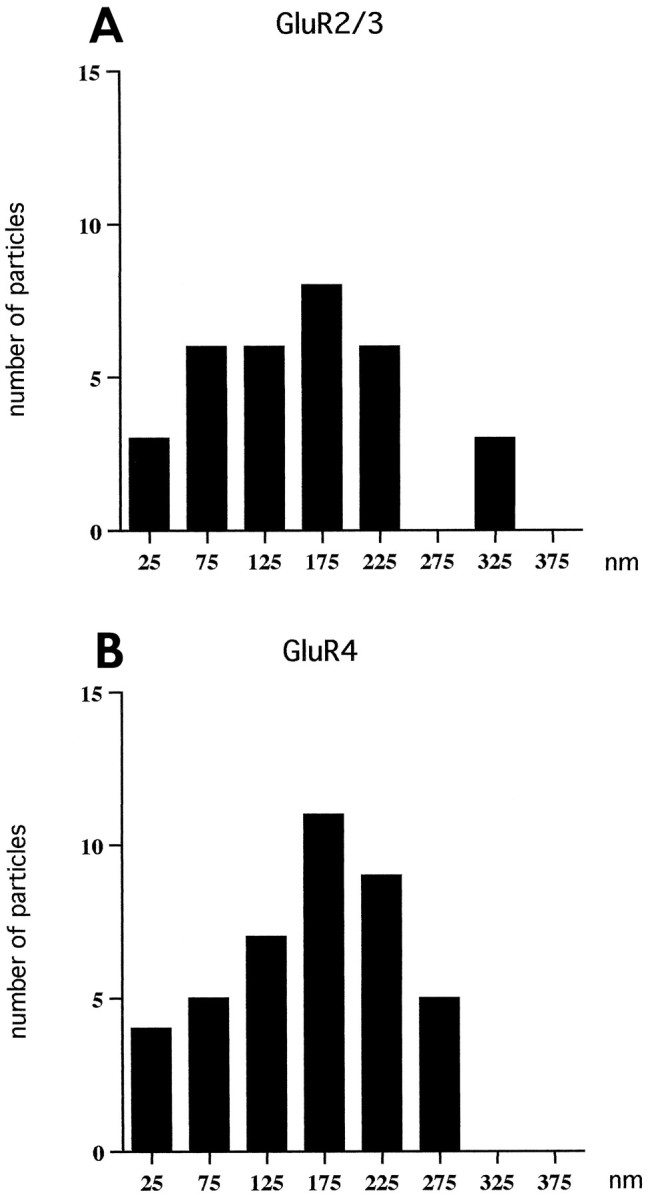
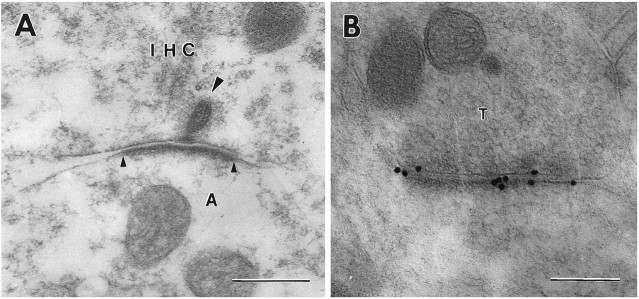
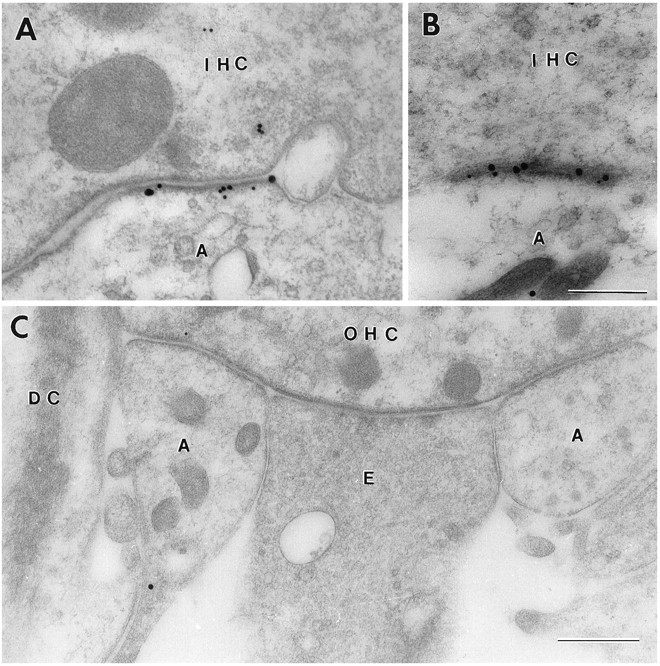
Sensitive and high-resolution immunocytochemical procedures were used to investigate the spatial organization of AMPA receptor subunits (GluR1-4) at the synapse between the inner hair cells and the afferent dendrites in the rat organ of Corti. This is a synapse with special functional properties and with a presynaptic dense body that defines the center of the synapse and facilitates its morphometric analysis. A quantitative postembedding immunocytochemical analysis was performed on specimens that had been embedded in a metachrylate resin at low temperature after freeze substitution. Single- and double-labeling procedures indicated that GluR2/3 and GluR4 subunits were colocalized throughout the postsynaptic density, with a maximum distance of 300 nm from the presynaptic body and with higher concentrations peripherally than centrally. No receptor immunolabeling was found at extrasynaptic membranes, but some GluR4 subunits appeared to be expressed presynaptically. The synapses between outer hair cells and afferent dendrites were devoid of labeling. The present data indicate that AMPA receptor subunits are inserted into the postsynaptic membrane in a very precise manner and that their density increases on moving away from the center of the synapse.
Figures

References
-
- Altschuler RA, Sheridan CE, Horn JW, Wenthold RJ. Immunocytochemical localization of glutamate immunoreactivity in the guinea pig cochlea. Hear Res. 1989;42:167–174. - PubMed
-
- Anniko M, Lundquist P-G. Temporal bone morphology after systemic arterial perfusion or intralabyrinthine in situ immersion. 1. Hair cells of the vestibular organs and the cochlea. Micron. 1980;11:73–83.
-
- Baude A, Nusser Z, Molnár E, McIlhinney RAJ, Somogyi P. High-resolution immunogold localization of AMPA type glutamate receptor subunits at synaptic and non-synaptic sites in rat hippocampus. Neuroscience. 1995;69:1031–1055. - PubMed
-
- Bennett JA, Dingledine R. Topology profile for a glutamate receptor: three transmembrane domains and a channel-lining reentrant membrane loop. Neuron. 1995;14:373–384. - PubMed
-
- Boulter J, Hollmann M, O’Shea-Greenfield A, Hartley M, Deneris E, Maron C, Heinemann S. Molecular cloning and functional expression of glutamate receptor subunit genes. Science. 1990;249:1033–1037. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources