

Mechanisms of H+ and Na+ changes induced by glutamate, kainate, and D-aspartate in rat hippocampal astrocytes
- PMID: 8757252
- PMCID: PMC6578898
- DOI: 10.1523/JNEUROSCI.16-17-05393.1996
Mechanisms of H+ and Na+ changes induced by glutamate, kainate, and D-aspartate in rat hippocampal astrocytes
Abstract
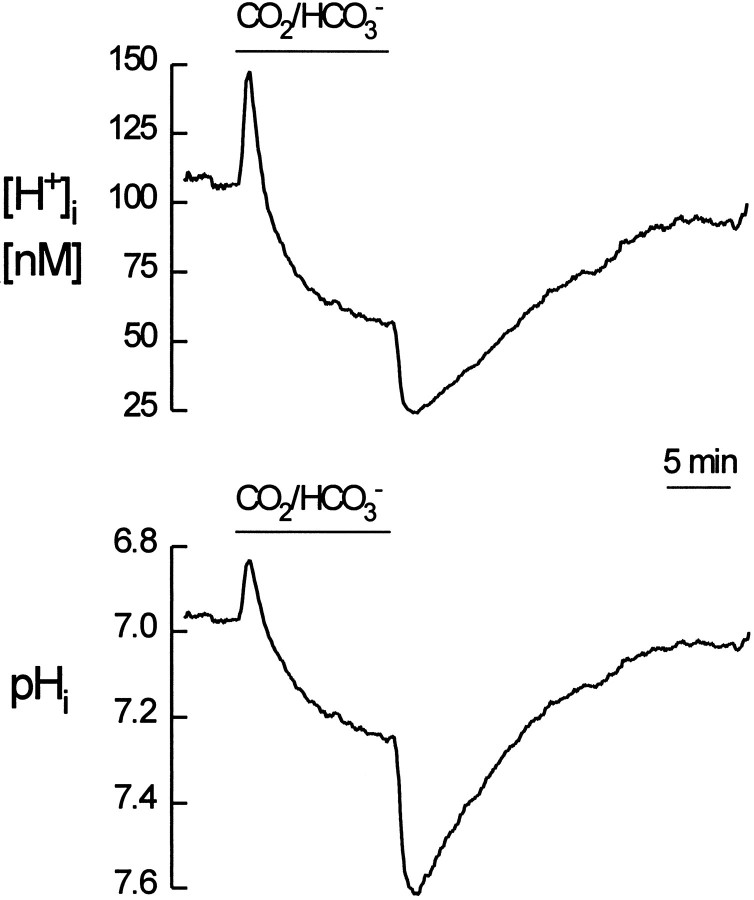
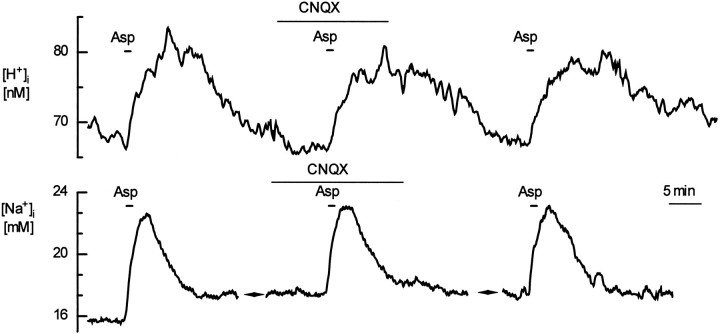
The excitatory transmitter glutamate (Glu), and its analogs kainate (KA), and D-aspartate (D-Asp) produce significant pH changes in glial cells. Transmitter-induced pH changes in glial cells, generating changes in extracellular pH, may represent a special form of neuronal-glial interaction. We investigated the mechanisms underlying these changes in intracellular H+ concentration ([H+]i) in cultured rat hippocampal astrocytes and studied their correlation with increases in intracellular Na+ concentration ([Na+]i), using fluorescence ratio imaging with 2',7'-bis(carboxyethyl)-5,6-carboxyfluorescein (BCECF) or sodium-binding benzofuran isophthalate (SBFI). Glu, KA, or D-Asp evoked increases in [Na+]i; Glu or D-Asp produced parallel acidifications. KA, in contrast, evoked biphasic changes in [H+]i, alkaline followed by acid shifts, which were unaltered after Ca2+ removal and persisted in 0 CI(-)-saline, but were greatly reduced in CO2/HCO3(-)-free or Na(+)-free saline, or during 4,4'-diisothiocyanato-stilbene-2,2'-disulphonic acid (DIDS) application. The non-NMDA receptor antagonist 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX) blocked KA-evoked changes in [H+]i and [Na+]i, indicating that they were receptor-ionophore mediated. In contrast, CNQX increased the [H+]i change and decreased the [Na+]i change induced by Glu. D-Asp, which is transported but does not act at Glu receptors, induced [H+]i and [Na+]i changes that were virtually unaltered by CNQX. Our study indicates that [Na+]i increases are not primarily responsible for Glu- or KA-induced acidifications in astrocytes. Instead, intracellular acidifications evoked by Glu or D-Asp are mainly caused by transmembrane movement of acid equivalents associated with Glu/Asp-uptake into astrocytes. KA-evoked biphasic [H+]i changes, in contrast, are probably attributable to transmembrane ion movements mediated by inward, followed by outward, electrogenic Na+/HCO3- cotransport, reflecting KA-induced biphasic membrane potential changes.
Figures










References
-
- Amato A, Ballerini L, Attwell D. Intracellular pH changes produced by glutamate uptake in rat hippocampal slices. J Neurophysiol. 1994;72:1686–1696. - PubMed
-
- Aronson PS. Kinetic properties of the plasma membrane Na+-H+ exchanger. Annu Rev Physiol. 1985;47:545–560. - PubMed
-
- Backus KH, Kettenmann H, Schachner M. Pharmacological characterization of the glutamate receptor in cultured astrocytes. J Neurosci Res. 1989;22:274–282. - PubMed
-
- Ballanyi K, Kettenmann H. Intracellular Na+activity in cultured mouse oligodendrocytes. J Neurosci Res. 1990;26:455–460. - PubMed
-
- Ballanyi K, Schlue WR, Dorner R. Glutamate and kainate increase intracellular sodium activity in leech neuropile glial cells. Glia. 1989;2:51–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous