

Use of 19F NMR to probe protein structure and conformational changes
- PMID: 8800468
- PMCID: PMC2899692
- DOI: 10.1146/annurev.bb.25.060196.001115
Use of 19F NMR to probe protein structure and conformational changes
Abstract
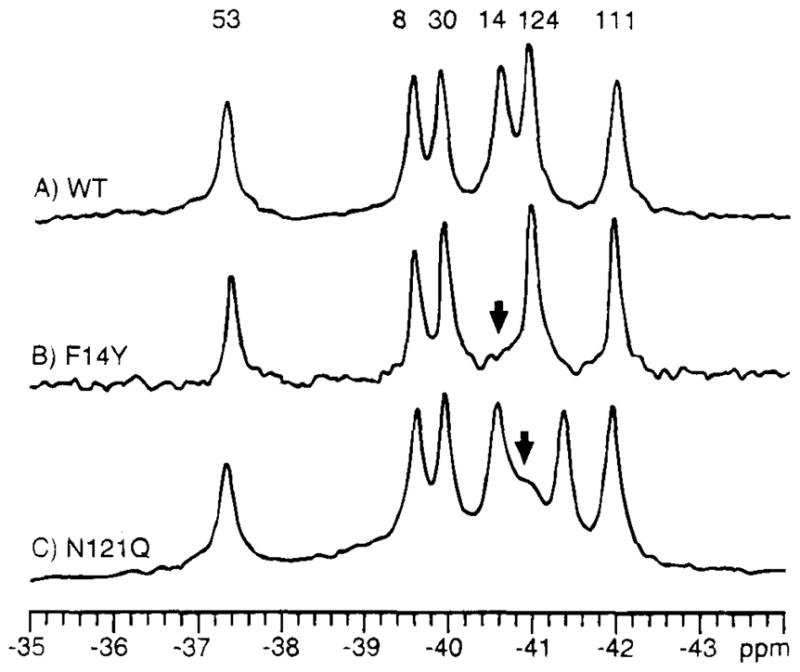
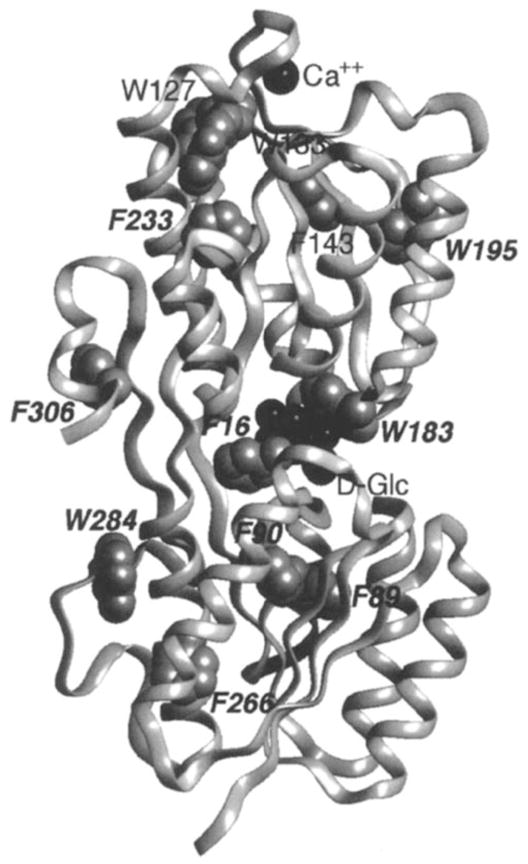
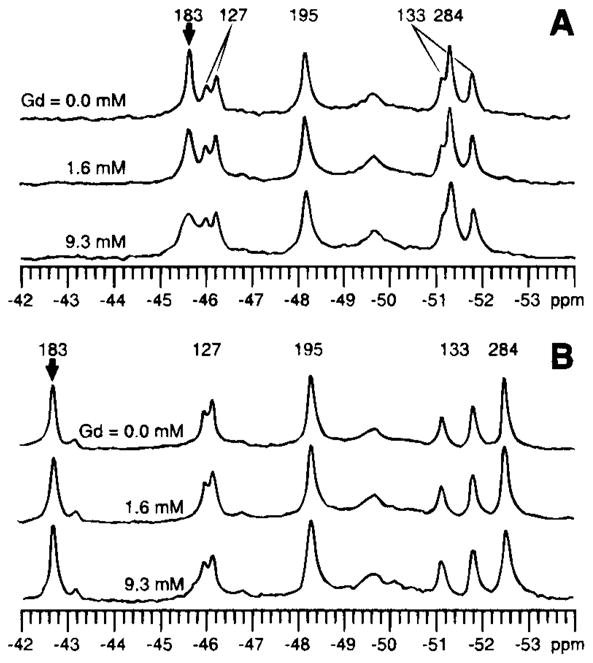
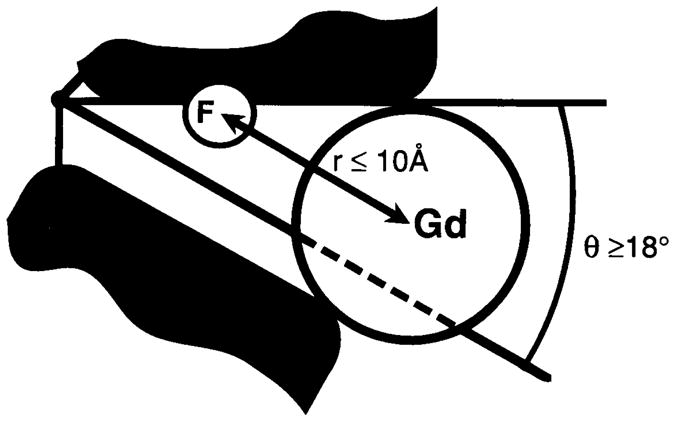
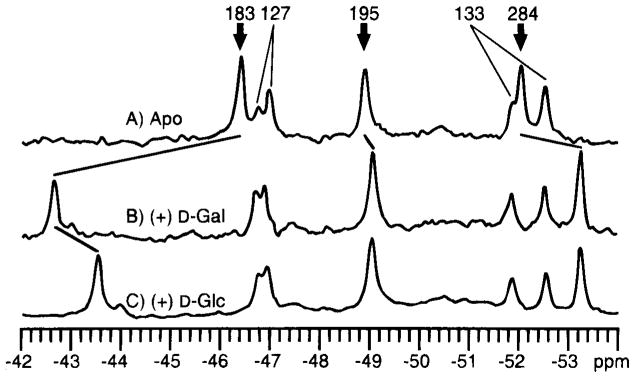
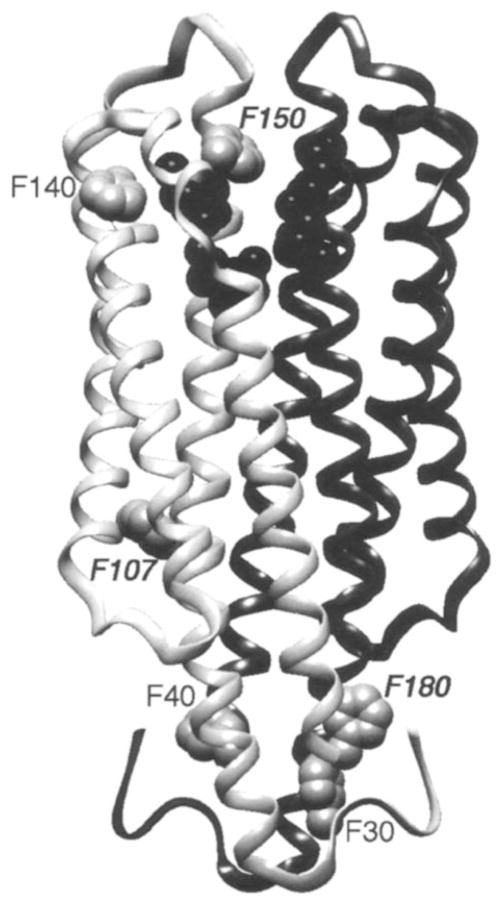
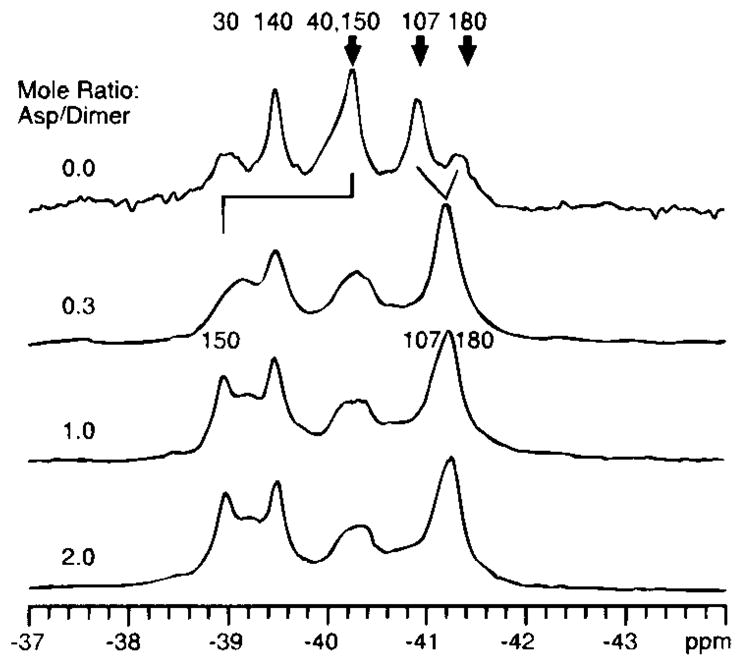
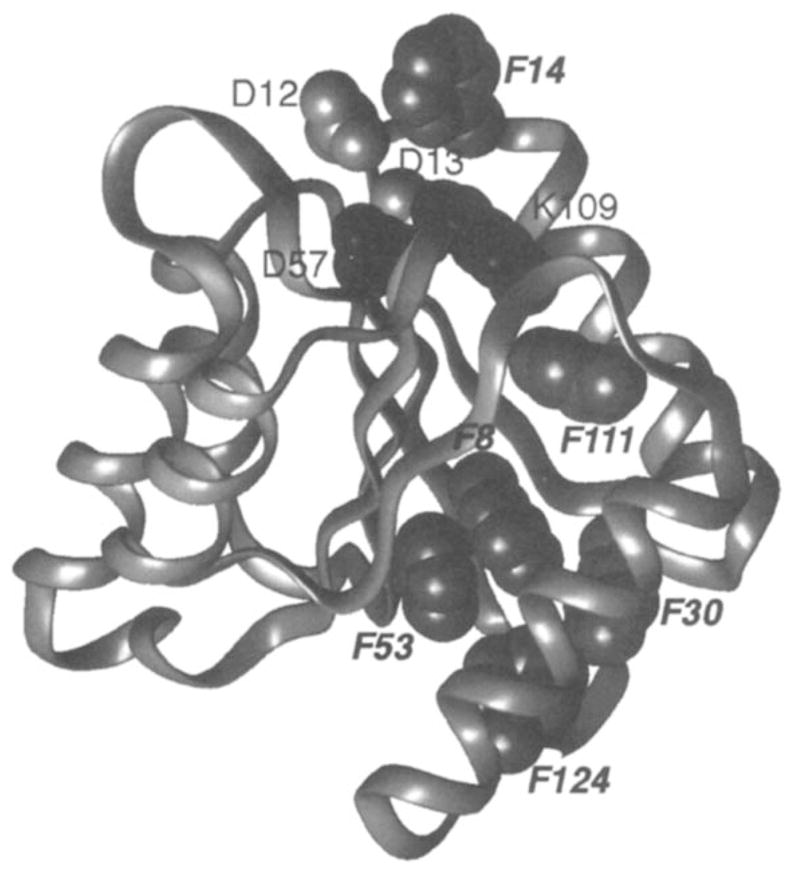
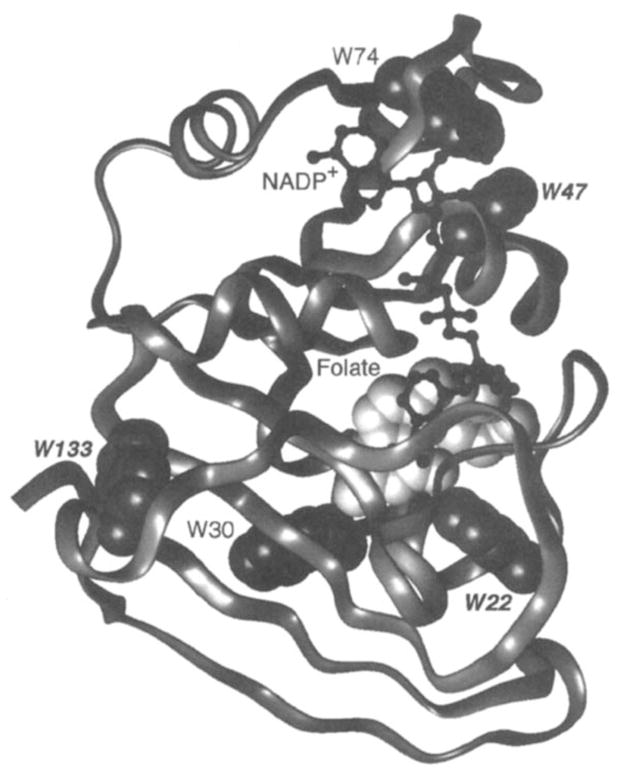
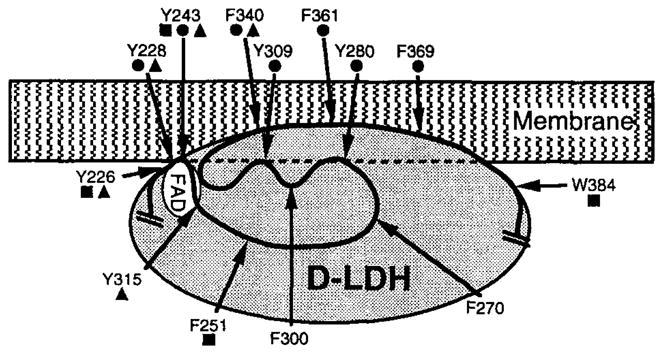
19F NMR has proven to be a powerful technique in the study of protein structure and dynamics because the 19F nucleus is easily incorporated at specific labeling sites, where it provides a relatively nonperturbing yet sensitive probe with no background signals. Recent applications of 19F NMR in mapping out structural and functional features of proteins, including the galactose-binding protein, the transmembrane aspartate receptor, the CheY protein, dihydrofolate reductase, elongation factor-Tu, and D-lactose dehydrogenase, illustrate the utility of 19F NMR in the analysis of protein conformational states even in molecules too large or unstable for full NMR structure determination. These studies rely on the fact that the chemical shift of 19F is extremely sensitive to changes in the local conformational environment, including van der Waals packing interactions and local electrostatic fields. Additional information is provided by solvent-induced isotope shifts or line broadening of the 19F resonance by aqueous and membrane-bound paramagnetic probes, which may reveal the proximity of a 19F label to bulk solvent or a biological membrane. Finally, the effect of exchanging conformations on the 19F resonance can directly determine the kinetic parameters of the conformational transition.
Figures
References
-
- Abeles RH, Alston TA. Enzyme inhibition by fluoro compounds. J Biol Chem. 1990;265:16705–8. - PubMed
-
- Arseniev AS, Kuryatov AB, Tsetlin VI, Bystrov VF, Ivanov VT, Orchinnikov YA. 19F NMR study of 5-fluorotrytophan-labeled bacteriorhodopsin. FEBS Lett. 1987;213:283–88.
-
- Augspurger J, Pearson JG, Oldfield E, Dykstra CE, Park KD, Schwartz D. Chemical-shift ranges in proteins. J Magn Reson. 1992;100:342–57.
-
- Bellsolell L, Prieto J, Serrano L, Coll M. Magnesium binding to the bacterial chemotaxis protein CheY results in large conformational changes involving its functional surface. J Mol Biol. 1994;238:489–95. - PubMed
-
- Biemann H-P, Koshland DE., Jr Aspartate receptors of Escherichia coli and Salmonella typhimurium bind ligand with negative and half-of-sites cooperativity. Biochemistry. 1994;33:629–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources