

Excitatory GABA responses in embryonic and neonatal cortical slices demonstrated by gramicidin perforated-patch recordings and calcium imaging
- PMID: 8815920
- PMCID: PMC6578913
- DOI: 10.1523/JNEUROSCI.16-20-06414.1996
Excitatory GABA responses in embryonic and neonatal cortical slices demonstrated by gramicidin perforated-patch recordings and calcium imaging
Abstract
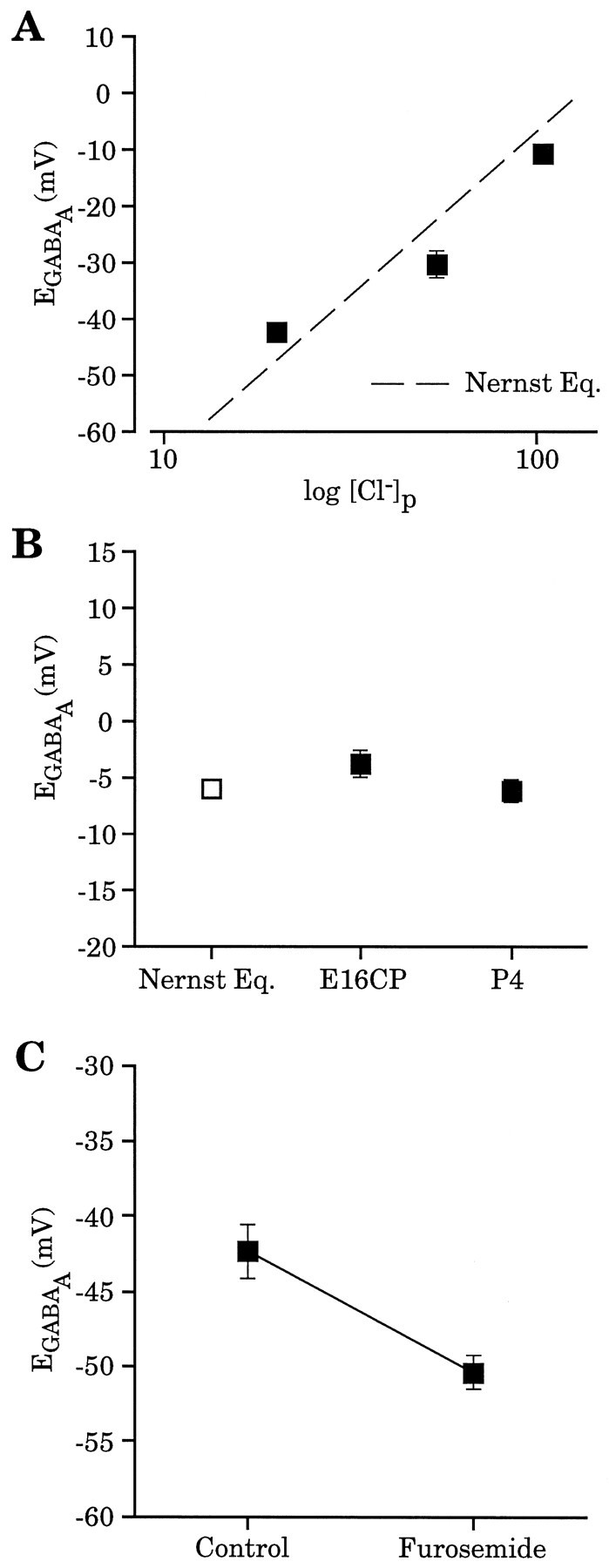
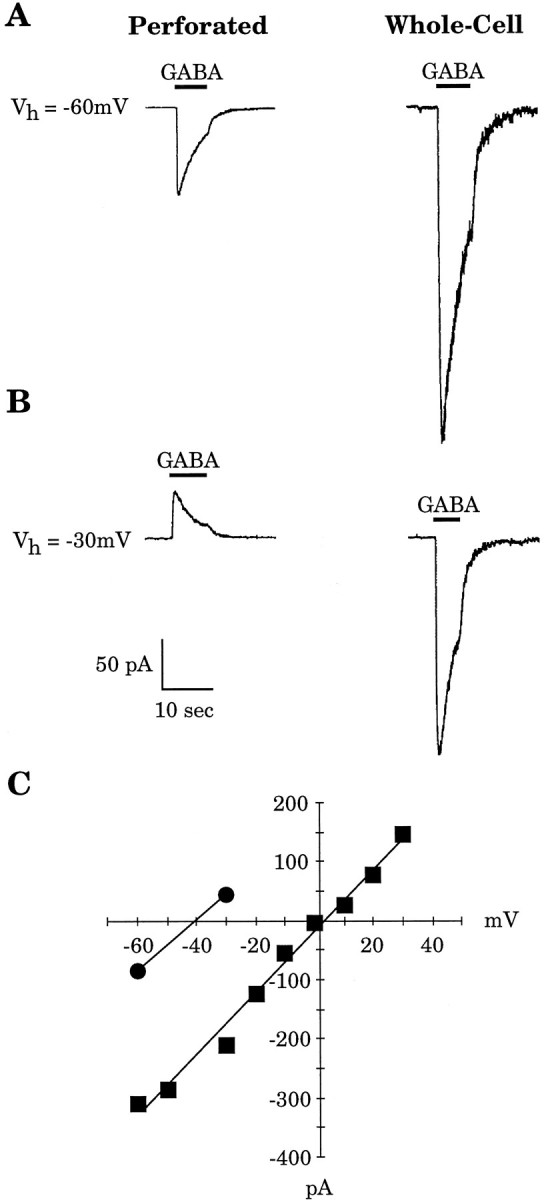
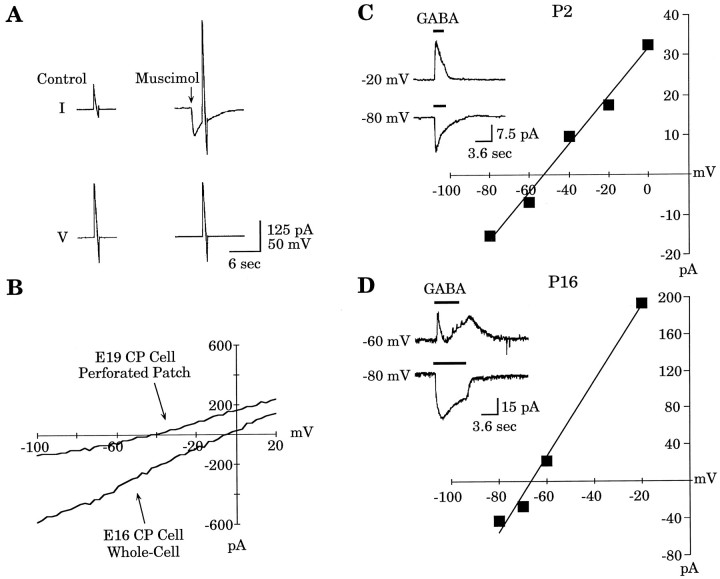
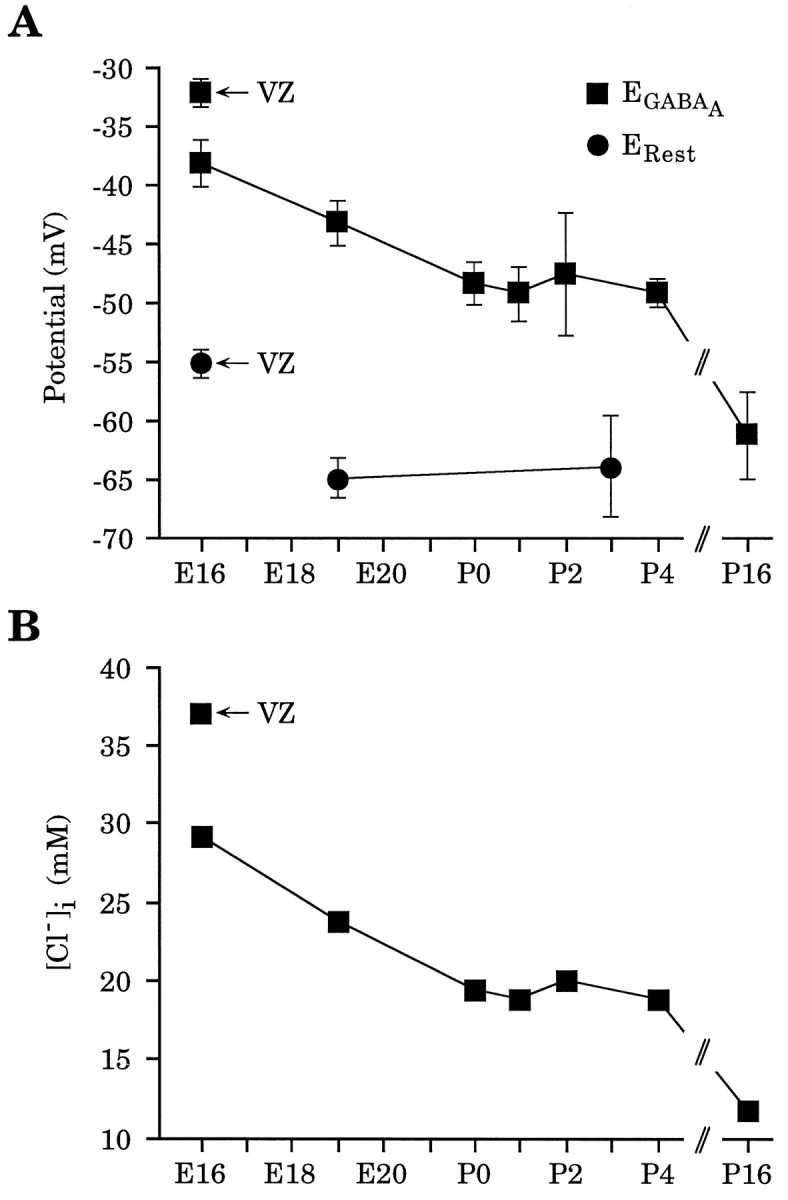
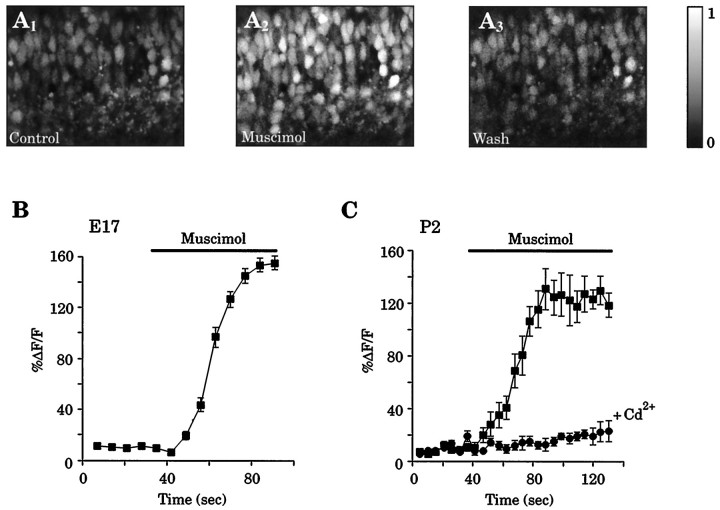
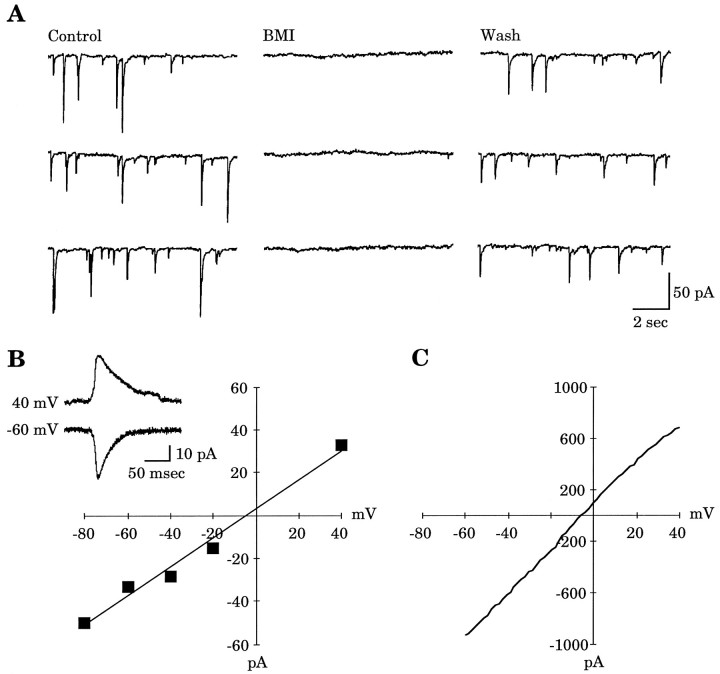
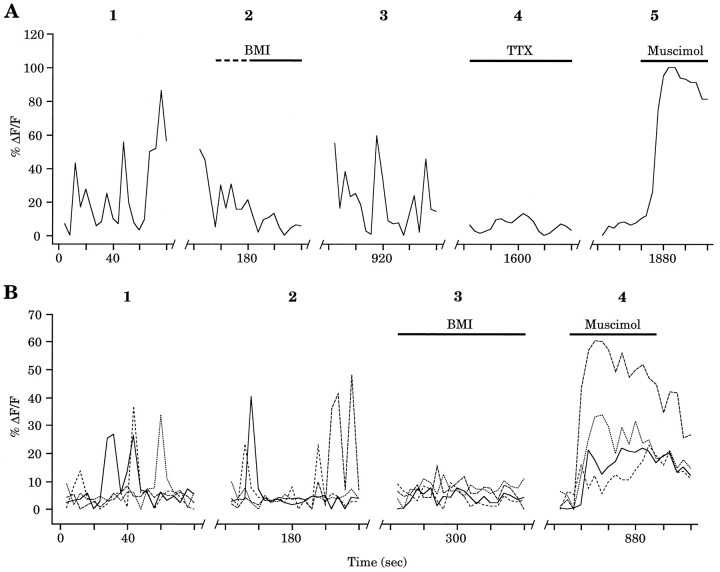
Gramicidin perforated-patch-clamp recordings in brain slices were used to obtain an accurate assessment of the developmental change in the GABAA receptor reversal potential (EGABAA) in embryonic and early postnatal rat neocortical cells including neuroepithelial precursor cells, cortical plate neurons, and postnatal neocortical neurons. Our results demonstrate that there is a progressive negative shift in EGABAA with the most positive values found in the youngest cortical precursor cells. At the early stages of neocortical development, EGABAA is determined by the chloride (Cl-) gradient, and the internal chloride concentration ([Cl-]i) decreases with development. EGABAA is positive to the resting potential, indicating that GABA serves to depolarize developing neocortical cells. Consistent with this conclusion, GABAA receptor activation with muscimol was found-to increase the internal calcium concentration ([Ca2+]i) in both embryonic and early postnatal neocortical cells through the activation of voltage-gated calcium channels (VGCCs). Postnatal cells exhibit spontaneous postsynaptic synaptic currents, which are eliminated by bicuculline methiodide (BMI) but not glutamate receptor antagonists and reverse at the Cl- equilibrium potential. Likewise, brief spontaneous increases in [Ca2+]i, sensitive to BMI and TTX, are observed at the same ages, suggesting that endogenous synaptic GABAA receptor activation can depolarize cells and activate VGCCs. These results suggest that GABAA receptor-mediated depolarization may influence early neocortical developmental events, including neurogenesis and synaptogenesis, through the activation of Ca(2+)-dependent signal transduction pathways.
Figures
References
-
- Abe Y, Furukawa K, Itoyama Y, Akaike N. Glycine response in acutely dissociated ventromedial hypothalamic neuron of the rat: new approach with gramicidin perforated patch-clamp technique. J Neurophysiol. 1994;72:1530–1537. - PubMed
-
- Araki T, Kiyama H, Tohyama M. GABAA receptor subunit messenger RNAs show differential expression during cortical development in the rat brain. Neuroscience. 1992;51:583–591. - PubMed
-
- Barbin G, Pollard H, Gaiarsa JL, Ben-Ari Y. Involvement of GABAA receptors in the outgrowth of cultured hippocampal neurons. Neurosci Lett. 1993;152:150–154. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous