

Differential survival of Cajal-Retzius cells in organotypic cultures of hippocampus and neocortex
- PMID: 8824328
- PMCID: PMC6579265
- DOI: 10.1523/JNEUROSCI.16-21-06896.1996
Differential survival of Cajal-Retzius cells in organotypic cultures of hippocampus and neocortex
Abstract
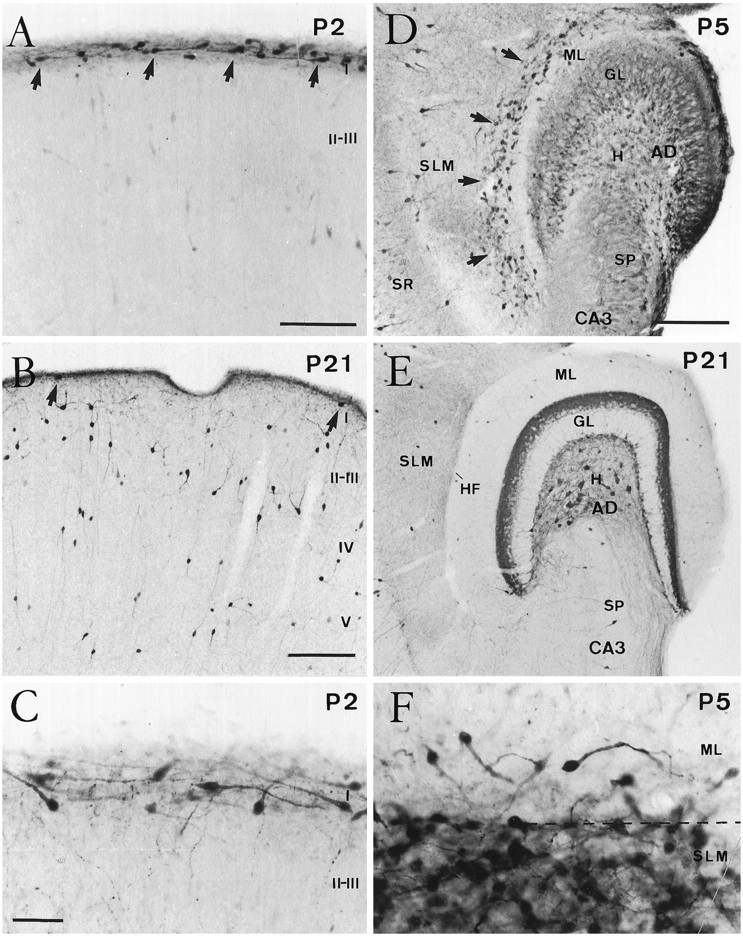
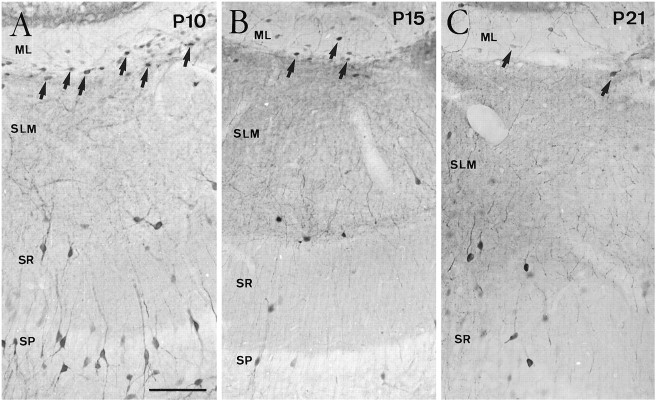
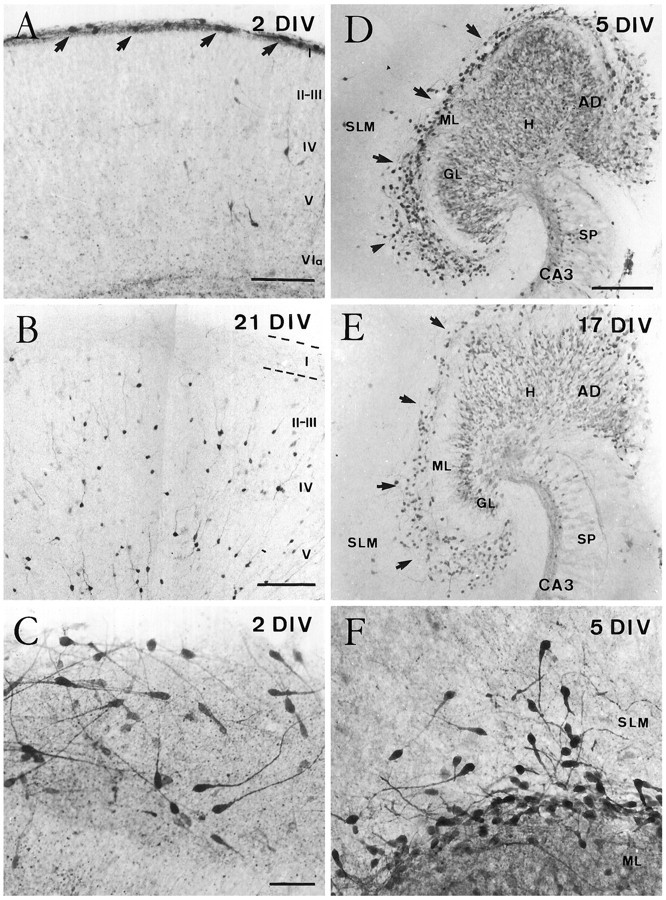
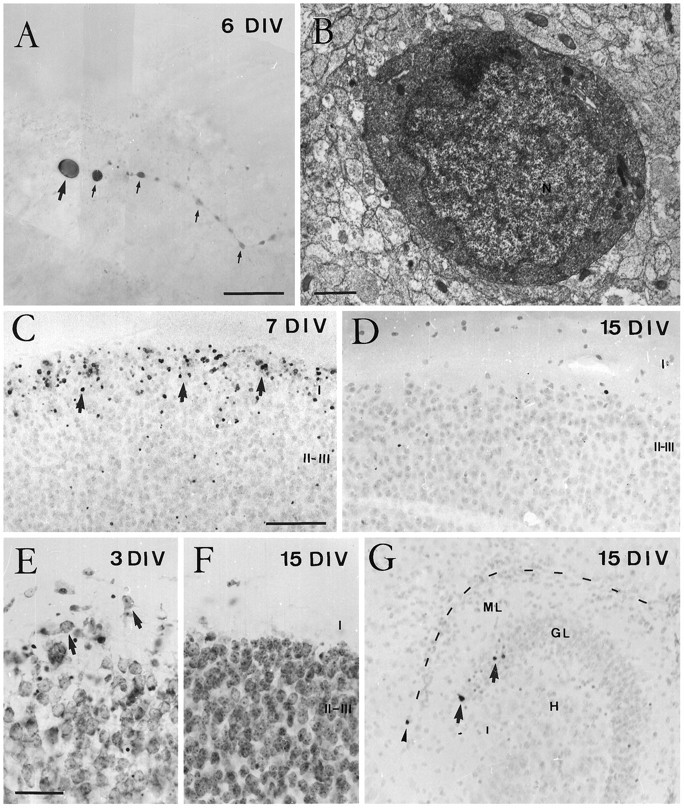
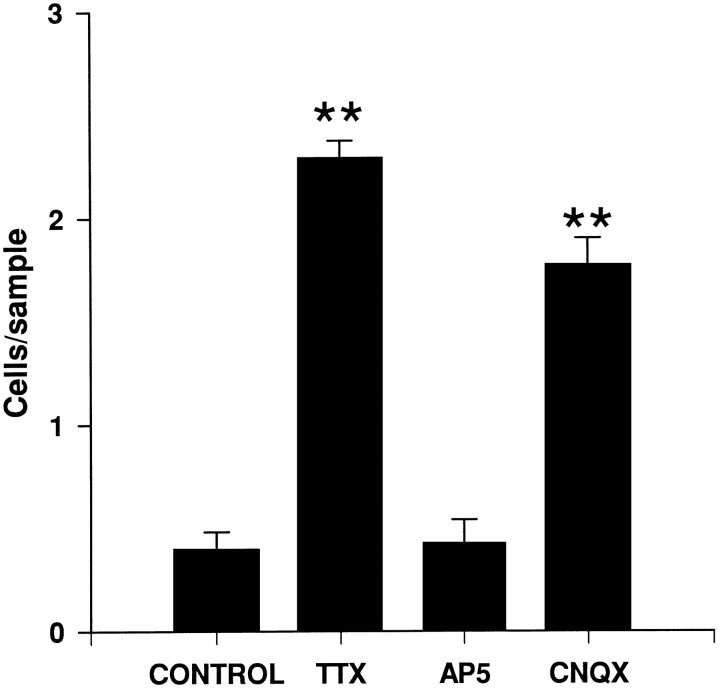
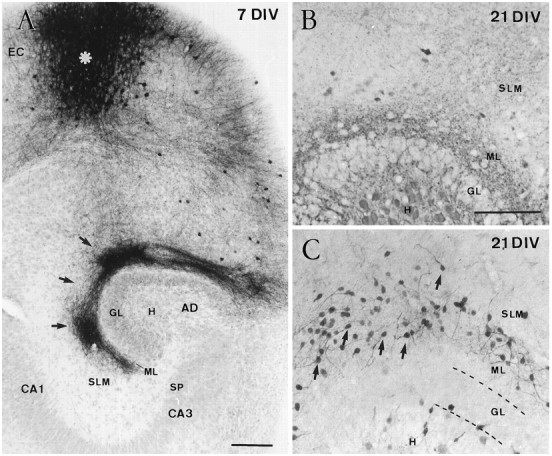
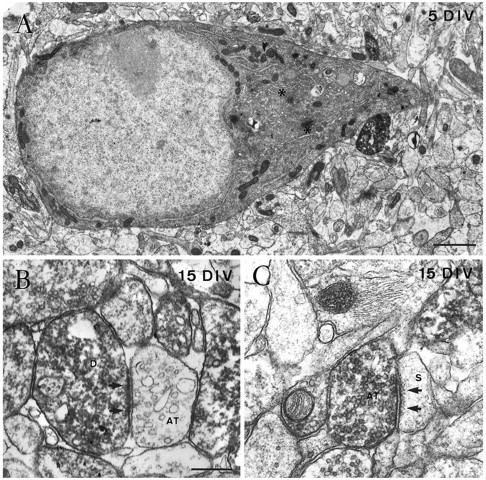
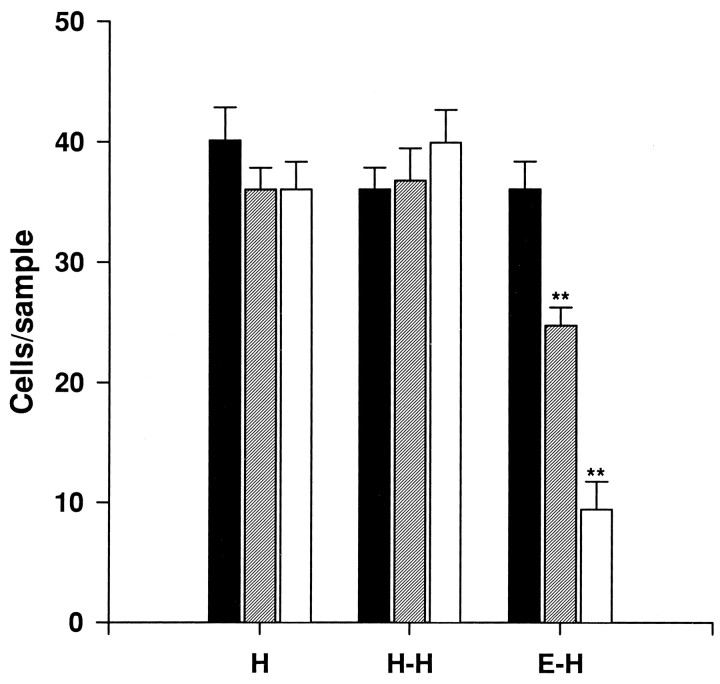
Cajal-Retzius (CR) cells are transient, pioneer neurons of layer I of the cortex that are believed to play essential roles in corticogenesis, e.g., in neuronal migration and synaptogenesis. Here we have used calretinin immunostaining to study the characteristics, survival, and fate of CR cells in single organotypic slice cultures of mouse neocortex and hippocampus deprived of their extrinsic afferents. In neocortical explants, CR cells were observed after 1-3 d in vitro (DIV), but they disappeared after 5-7 DIV, which is similar to their time of degeneration in vivo. The disappearance of CR cells in neocortical slices was prevented by incubation with tetrodotoxin and the glutamate receptor antagonist 6-cyano-7-nitroquinoxaline-2,3,-dione but not by 2-amino-5-phosphonopentanoic acid, suggesting that neuronal activity and non-NMDA glutamate receptors may trigger CR cell death in the neocortex. In contrast to the situation in vivo, in which many hippocampal CR cells disappear at approximately the third postnatal week, CR cells survived in single hippocampal cultures after long incubation times (31 DIV), with their morphology essentially unaltered. In contrast, fewer CR cells were found when hippocampal slices were cocultured with explants from the entorhinal cortex. Because CR cells are transient synaptic targets for entorhinohippocampal afferents, these findings suggest a role for entorhinal afferents in the degeneration of CR cells in the hippocampus. In conclusion, this study shows different survival properties of CR cells in organotypic slice cultures of hippocampus and neocortex, and it suggests that different mechanisms are involved in the regulation of the process of naturally occurring CR cell death in the two cortical regions.
Figures
References
-
- Abercrombie M. Estimation of nuclear population from microtome section. Anat Rec. 1946;94:297–247. - PubMed
-
- Al-Ghoul WM, Miller MV. Transient expression of Alz-50 immunoreactivity in developing rat neocortex: a marker for naturally neuronal death? Brain Res. 1989;481:361–367. - PubMed
-
- Allendoerfer KL, Shatz CJ. The subplate, a transient neocortical structure: its role in the development of connections between thalamus and cortex. Annu Rev Neurosci. 1994;17:185–218. - PubMed
-
- Bayer SA, Altman J. Development of layer I and the subplate in rat neocortex. Exp Neurol. 1990;107:48–62. - PubMed
-
- Bayer SA, Altman J (1991) Neocortical development. New York: Raven.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources