

Ovarian steroid regulation of tryptophan hydroxylase mRNA expression in rhesus macaques
- PMID: 8824338
- PMCID: PMC6579247
- DOI: 10.1523/JNEUROSCI.16-21-07021.1996
Ovarian steroid regulation of tryptophan hydroxylase mRNA expression in rhesus macaques
Abstract
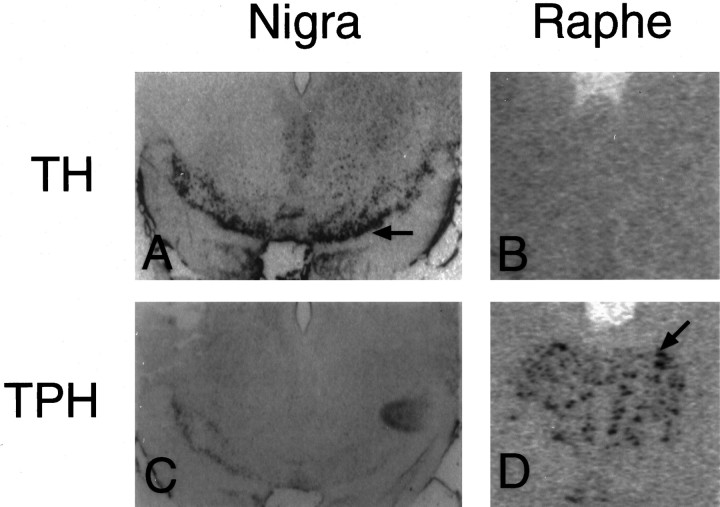
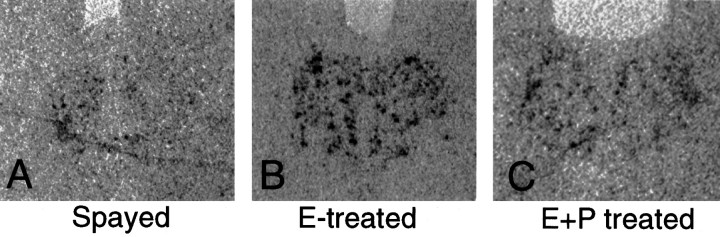
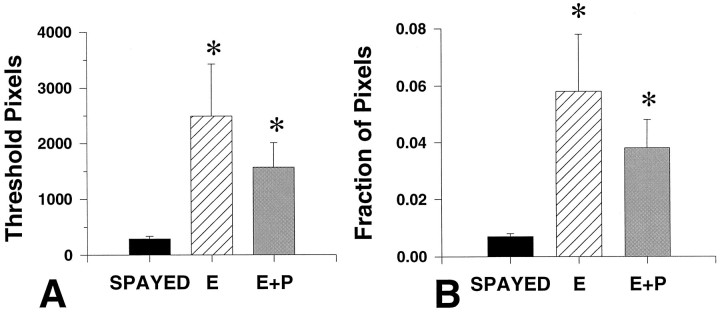
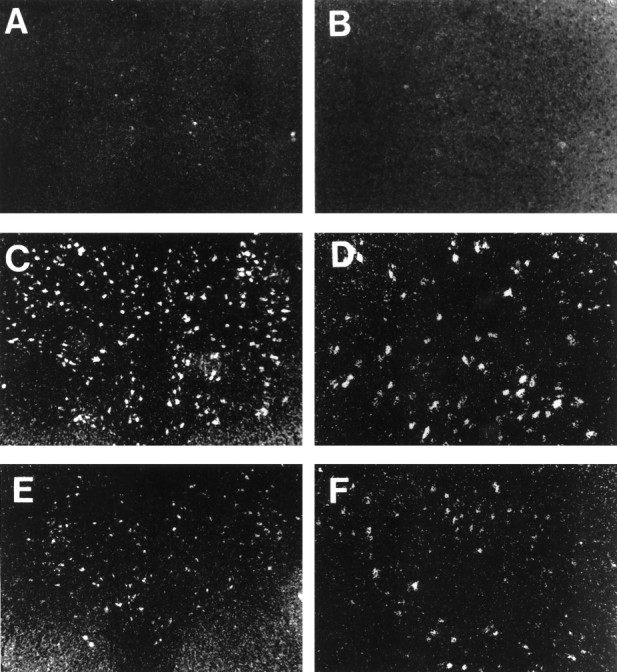
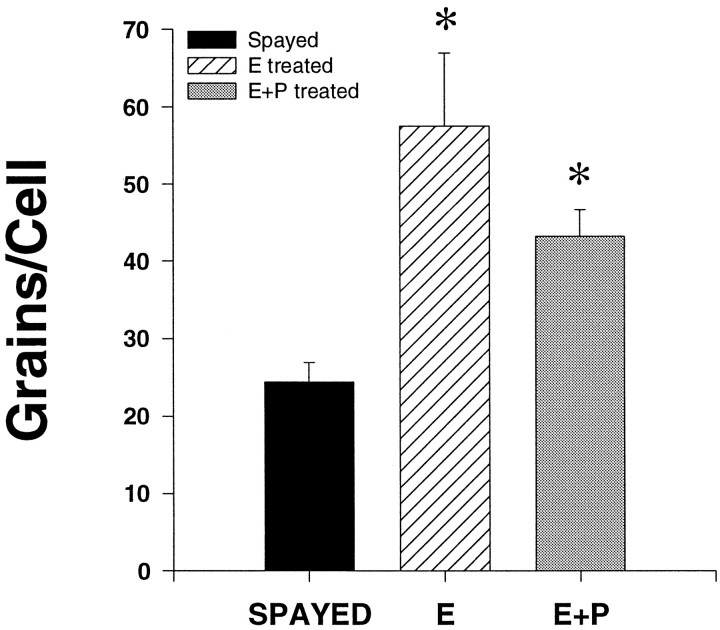
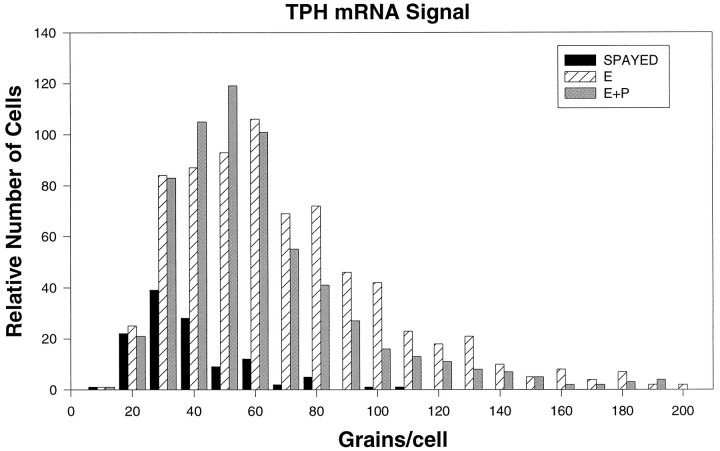
Progesterone (P) stimulates prolactin secretion through an unknown neural mechanism in estrogen (E)-primed female monkeys. Serotonin is a stimulatory neurotransmitter in prolactin regulation, and this laboratory has shown previously that E induces progestin receptors (PR) in serotonin neurons. Therefore, we questioned whether E and/or E+P increased serotonin neural function. The expression of mRNA for tryptophan hydroxylase (TPH) was examined in ovariectomized (spayed) control, E-treated (28 d), and E+P-treated monkeys (14 d E and 14 d E+P) using in situ hybridization and a 249 bp TPH cRNA probe generated with RT-PCR (n = 5 animals/group). Densitometric analysis of film autoradiographs revealed a ninefold increase in TPH mRNA in E-treated macaques compared to spayed animals (p < 0.05). With supplemental P treatment, TPH mRNA signal was increased fivefold over spayed animals (p < 0.05), but was not significantly different compared to E-treated animals. These results were verified by grain counts from photographic emulsion-coated slides. There were significantly higher single-cell levels of TPH mRNA in serotonergic neurons of the dorsal raphe in E- and E+P-treated groups (p < 0.05). These data indicate that E induces TPH gene expression in nonhuman primates and that the addition of P has little additive effect on TPH gene expression. Thus, the action of P on prolactin secretion is probably not mediated at the level of TPH gene transcription. However, because P increases raphe serotonin content in E-primed rodents, the possibility remains that P may have other actions on post-translational processing or enzyme activity.
Figures

References
-
- Anderson IM, Ware CJ, Da Roza Davis JM, Cowen PJ. Decreased 5-HT-mediated prolactin release in major depression. Br J Psychiatry. 1992;160:372–378. - PubMed
-
- Bethea CL. Colocalization of progestin receptors with serotonin in the raphe neurons of macaque. Neuroendocrinology. 1993;57:1–6. - PubMed
-
- Bethea CL. Regulation of progestin receptors in raphe neurons of steroid-treated monkeys. Neuroendocrinology. 1994;60:50–61. - PubMed
-
- Bethea CL, Papkoff H. Purification of monkey prolactin from culture medium: biochemical and immunological characterization. Proc Soc Exp Biol Med. 1986;182:22–33. - PubMed
-
- Bethea CL, Fahrenbach WH, Sprangers SA, Freesh F. Immunocytochemical localization of progestin receptors in monkey hypothalamus: effect of estrogen and progestin. Endocrinology. 1992;130:895–905. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials