

Direct physical interaction between DnaG primase and DnaB helicase of Escherichia coli is necessary for optimal synthesis of primer RNA
- PMID: 8917517
- PMCID: PMC24018
- DOI: 10.1073/pnas.93.23.12902
Direct physical interaction between DnaG primase and DnaB helicase of Escherichia coli is necessary for optimal synthesis of primer RNA
Abstract
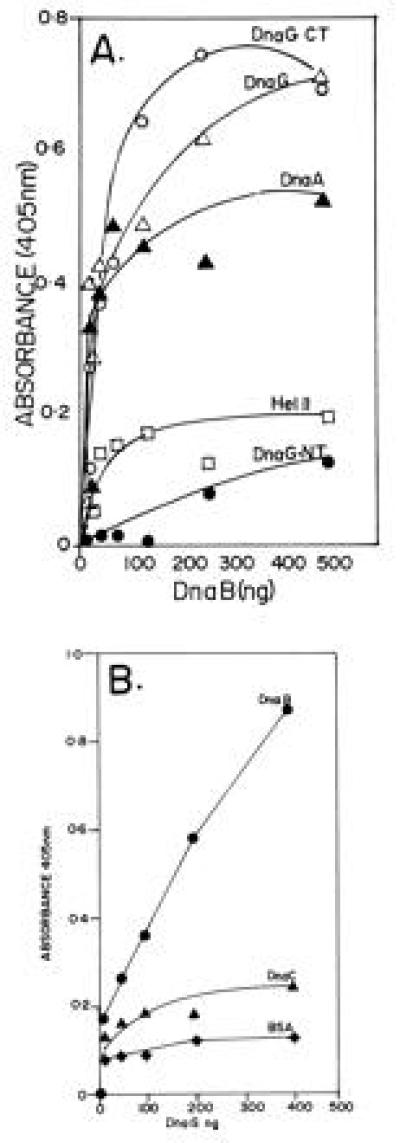
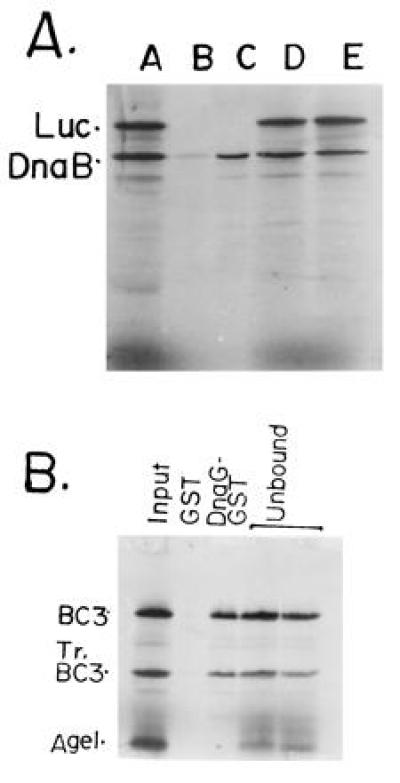
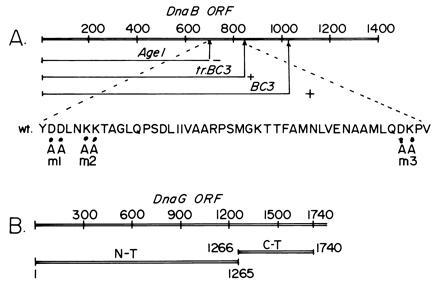
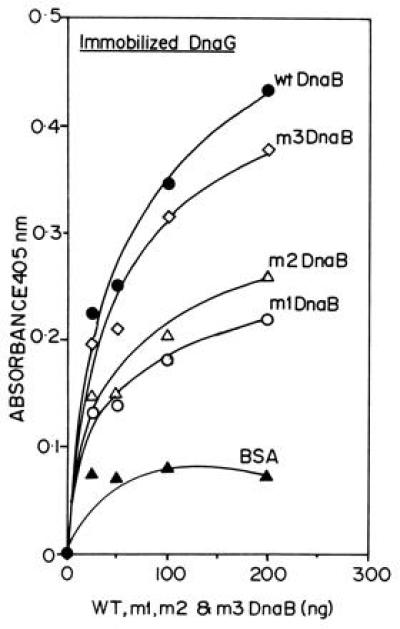
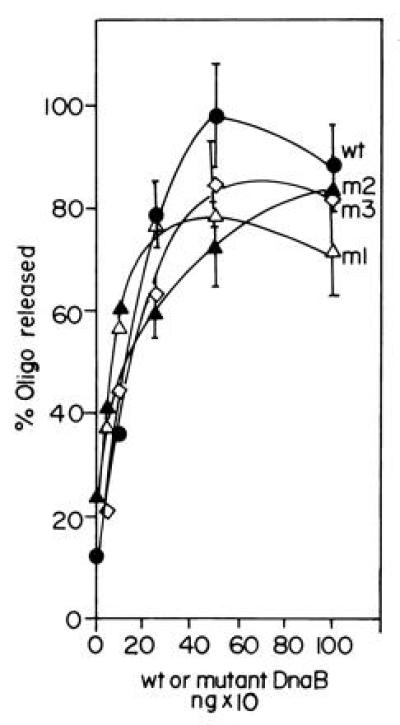
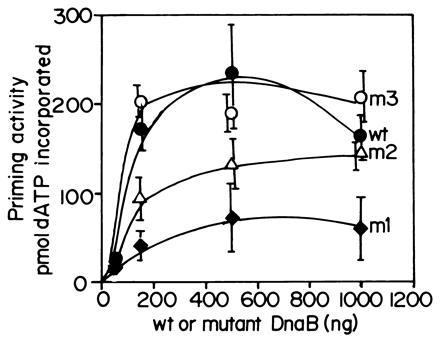
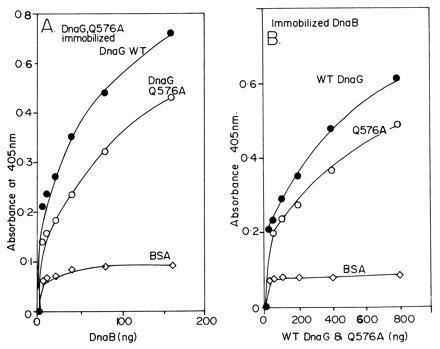
The primase DnaG of Escherichia coli requires the participation of the replicative helicase DnaB for optimal synthesis of primer RNA for lagging strand replication. However, previous studies had not determined whether the activation of the primase or its loading on the template was accomplished by a helicase-mediated structural alteration of the single-stranded DNA or by a direct physical interaction between the DnaB and the DnaG proteins. In this paper we present evidence supporting direct interaction between the two proteins. We have mapped the surfaces of interaction on both DnaG and DnaB and show further that mutations that reduce the physical interation also cause a significant reduction in primer synthesis. Thus, the physical interaction reported here appears to be physiologically significant.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
