

Interaction of measles virus glycoproteins with the surface of uninfected peripheral blood lymphocytes induces immunosuppression in vitro
- PMID: 8917567
- PMCID: PMC24069
- DOI: 10.1073/pnas.93.23.13194
Interaction of measles virus glycoproteins with the surface of uninfected peripheral blood lymphocytes induces immunosuppression in vitro
Abstract
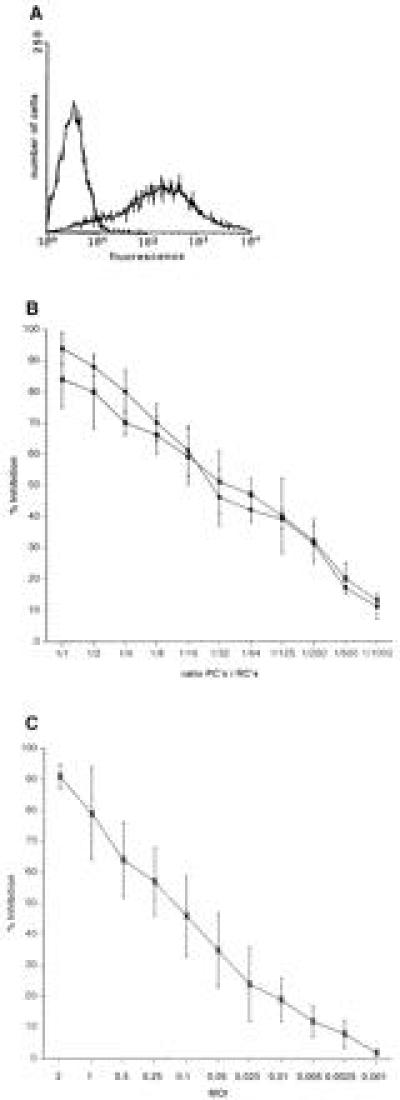
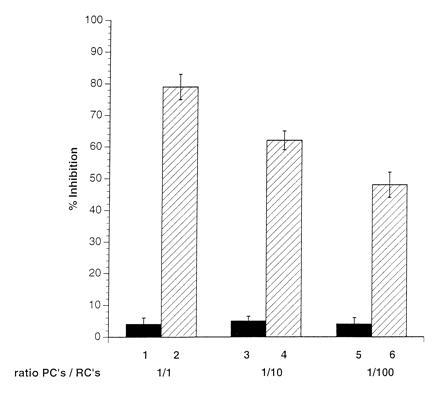
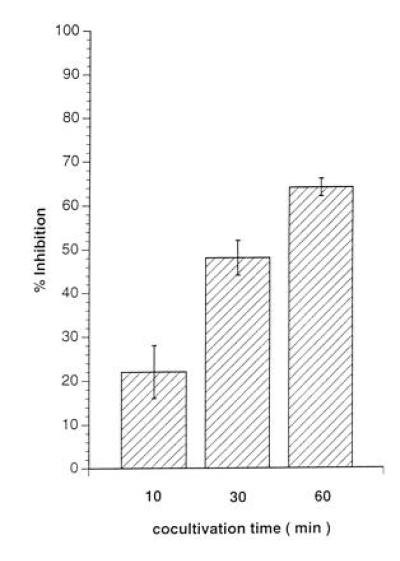
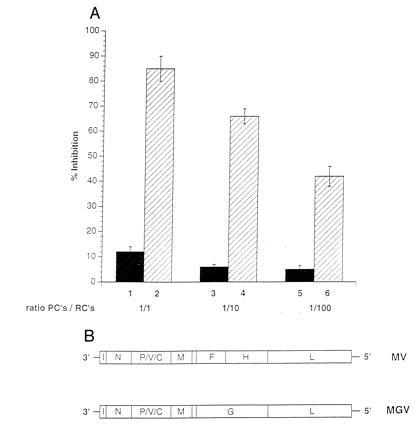
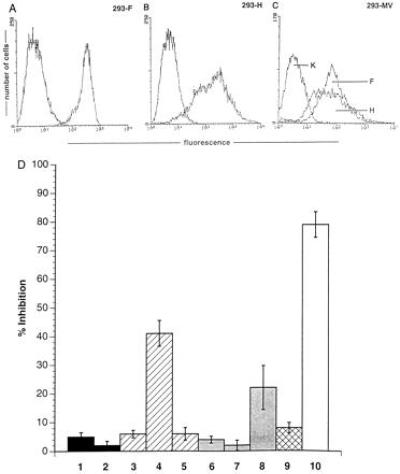
A marked suppression of immune function has long been recognized as a major cause of the high morbidity and mortality rate associated with acute measles. As a hallmark of measles virus (MV)-induced immunosuppression, peripheral blood lymphocytes (PBLs) isolated from patients exhibit a significantly reduced capacity to proliferate in response to mitogens, allogens, or recall antigens. In an in vitro system we show that proliferation of naive PBLs [responder cells (RCs)] in response to a variety of stimuli was significantly impaired after cocultivation with MV-infected, UV-irradiated autologous PBLs [presenter cells (PCs]. We further observed that a 50% reduction in proliferation of RCs could still be observed when the ratio of PC to RC was 1:100. The effect was completely abolished after physical separation of the two populations, which suggests that soluble factors were not involved. Proliferative inhibition of the RCs was observed after short cocultivation with MV-infected cells, which indicates that surface contact between one or more viral proteins and the RC population was required. We identified that the complex of both MV glycoproteins, F and H, is critically involved in triggering MV-induced suppression of mitogen-dependent proliferation, since the effect was not observed (i) using a recombinant MV in which F and H were replaced with vesicular stomatitis virus G or (ii) when either of these proteins was expressed alone. Coexpression of F and H, however, lead to a significant proliferative inhibition in the RC population. Our data indicate that a small number of MV-infected PBLs can induce a general nonresponsiveness in uninfected PBLs by surface contact, which may, in turn, account for the general suppression of immune responses observed in patients with acute measles.
Figures
Comment in
-
Virus-lymphoid cell interactions.Proc Natl Acad Sci U S A. 1996 Nov 12;93(23):12756-8. doi: 10.1073/pnas.93.23.12756. Proc Natl Acad Sci U S A. 1996. PMID: 8917490 Free PMC article. Review. No abstract available.
References
-
- Clements C J, Cutts F. Curr Top Microbiol Immunol. 1995;191:13–34. - PubMed
-
- Casali P, Nakamura M, McChesney M B. In: Virus-Induced Immunosuppression. Spector S, Bendinelli M, Friedman H, editors. New York: Plenum; 1989. pp. 345–373.
-
- Borrow P, Oldstone M B A. Curr Top Microbiol Immunol. 1995;191:85–99. - PubMed
-
- Smithwick E M, Berkovich S. In: Cellular Recognition. Smith R T, Good R A, editors. New York: Appleton Century Crafts; 1969. p. 131.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
