

Two alternatively spliced forms of the cGMP-gated channel alpha-subunit from cone photoreceptor are expressed in the chick pineal organ
- PMID: 8922401
- PMCID: PMC6579096
- DOI: 10.1523/JNEUROSCI.16-23-07458.1996
Two alternatively spliced forms of the cGMP-gated channel alpha-subunit from cone photoreceptor are expressed in the chick pineal organ
Abstract
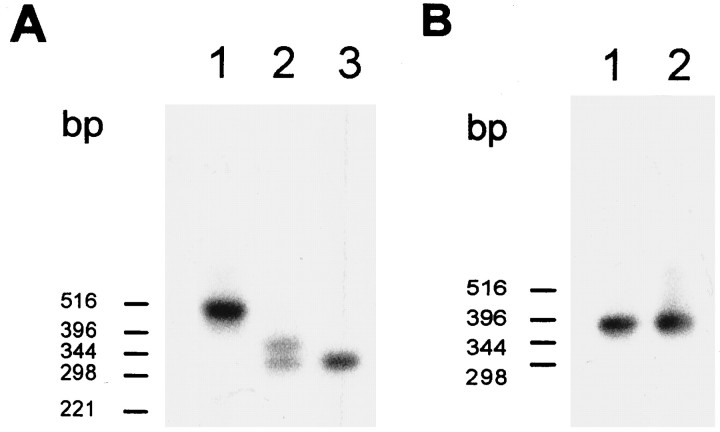
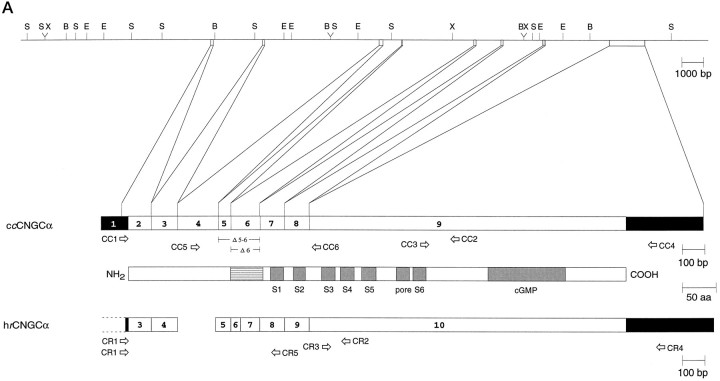
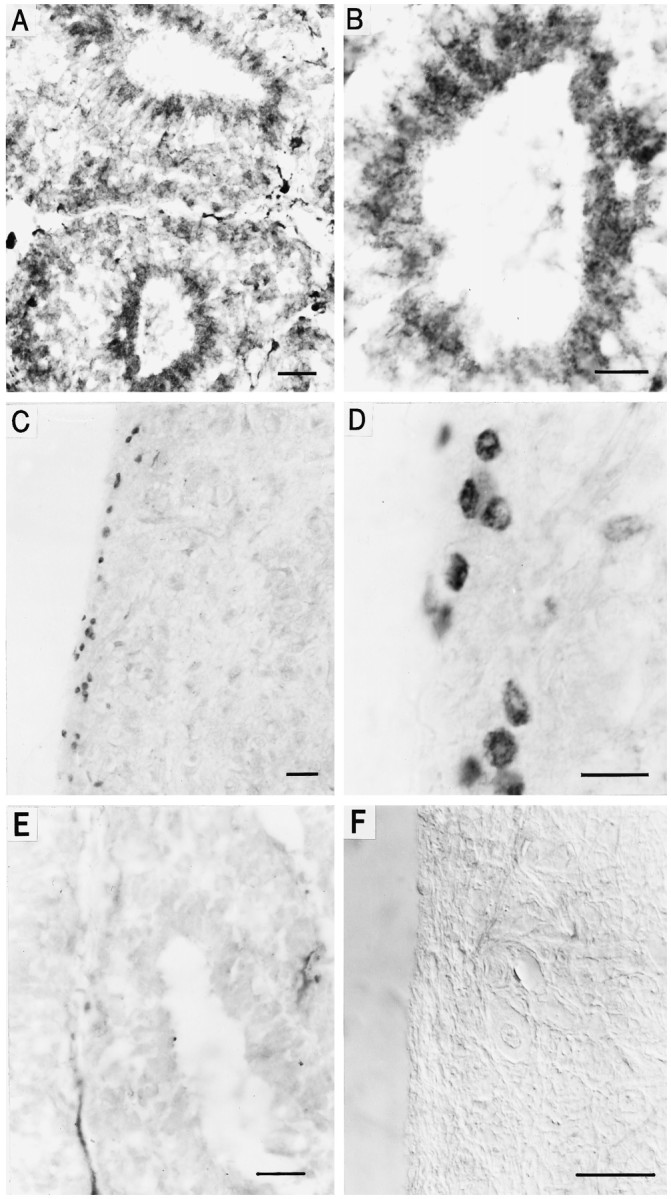
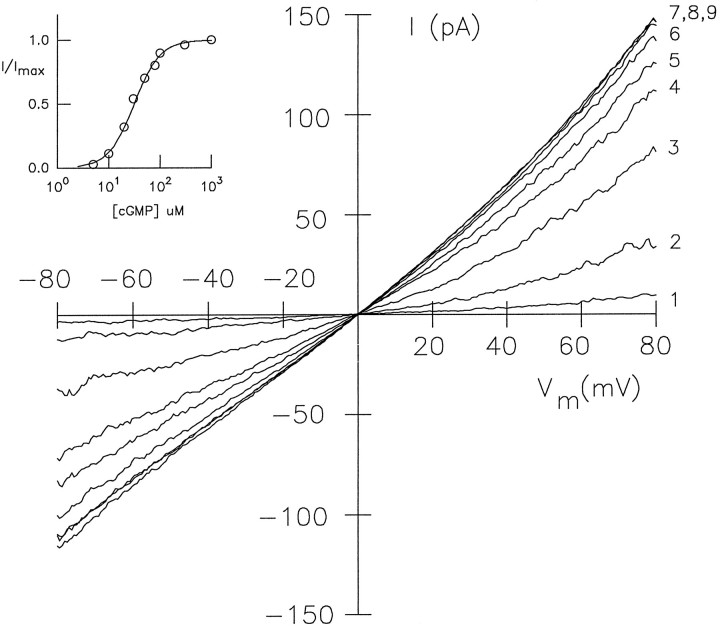
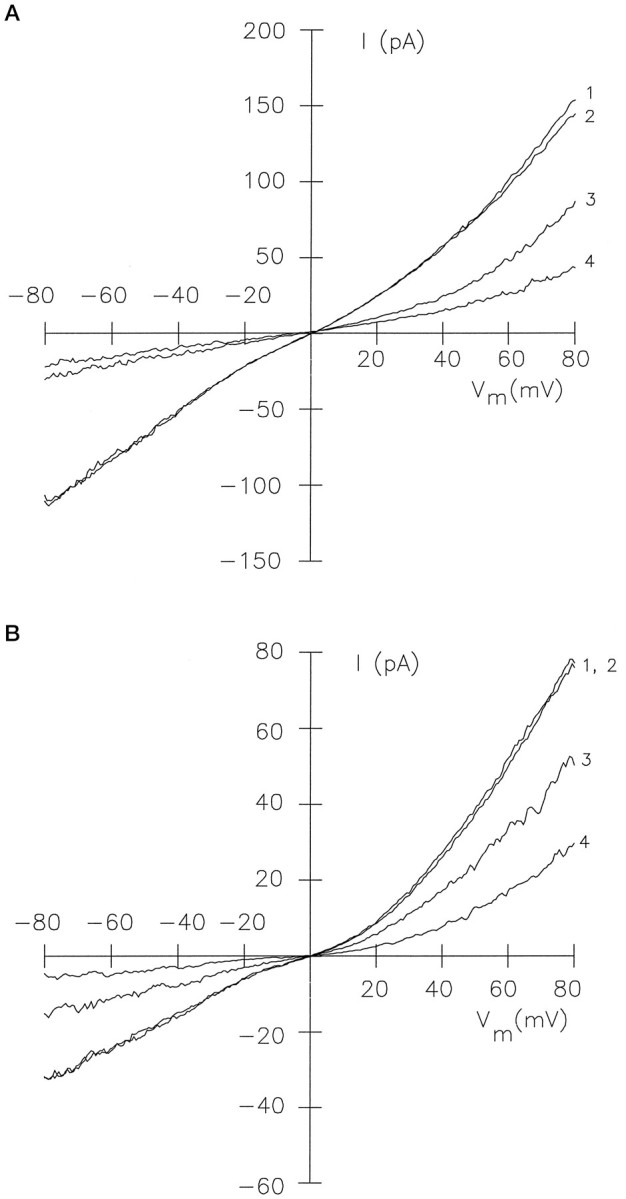
Light sensitivity of the pineal has been retained in most vertebrates, except mammals. Retinal photoreceptors and pinealocytes share common components of light-dependent signaling pathways. In particular, an ion channel gated by cGMP has been electrophysiologically identified in chick pinealocytes; however, the physiological function of a light-sensitive enzyme cascade is not known, and primary structures of only a few pineal components have been determined. By PCR analysis and cloning of the respective cDNA, we show that the chick pineal expresses the alpha-subunit of the cyclic nucleotide-gated (CNG) channel of rod photoreceptors and two short forms of the cone CNG channel. Analysis of the chick cone CNG channel gene reveals that these forms are produced by alternative splicing, which removes either one or two exons from the transcript. The shorter splice variant is functional when heterologously expressed, and it is approximately twofold more sensitive to activation by cGMP than the cone CNG channel. The chick cone CNG channel and the pineal splice form are both modulated by Ca2+/calmodulin (CaM). The CaM sensitivity might be mediated by a putative CaM-binding site in an N-terminal segment encoded by exon 4. This exon is missing in the gene for the rod CNG channel alpha-subunit. Pineal CNG channels are candidates for receptor-mediated Ca2+ entry into pinealocytes and may be an important element of signaling pathways that control the light response and secretion of the pineal hormone melatonin.
Figures


References
-
- Abe T, Nakabayashi H, Tamada H, Takagi T, Sakuragi S, Yamaki K, Shinohara T. Analysis of the human, bovine and rat 33 kDa proteins and cDNA in retina and pineal gland. Gene. 1990;91:209–215. - PubMed
-
- Abe T, Shinohara T. S-antigen from the rat retina and pineal gland have identical sequences. Exp Eye Res. 1990;51:111–112. - PubMed
-
- Bönigk W, Altenhofen W, Müller F, Dose A, Illing M, Molday RS, Kaupp UB. Rod and cone photoreceptor cells express distinct genes for cGMP-gated channels. Neuron. 1993;10:865–877. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous