

Direct measurement of AMPA receptor desensitization induced by glutamatergic synaptic transmission
- PMID: 8922405
- PMCID: PMC6579098
- DOI: 10.1523/JNEUROSCI.16-23-07496.1996
Direct measurement of AMPA receptor desensitization induced by glutamatergic synaptic transmission
Abstract
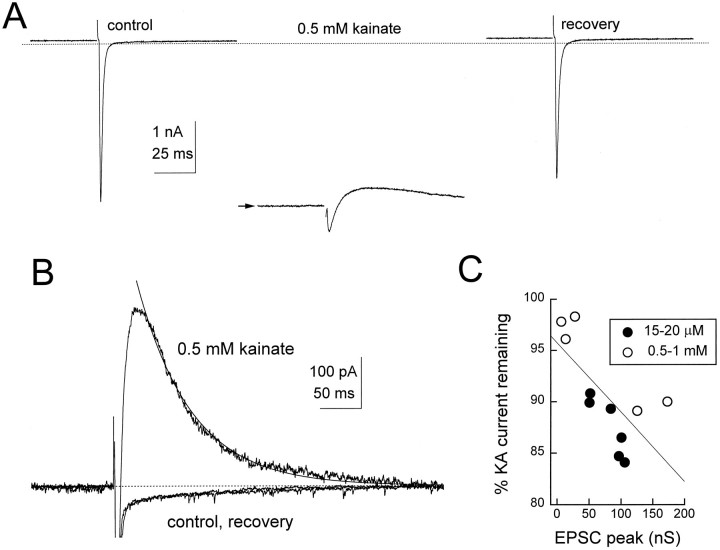
Although almost all ionotropic neurotransmitter receptors undergo desensitization, the onset and recovery of desensitization at a synapse have never been observed directly. We have found changes in postsynaptic AMPA receptor sensitivity in neurons of the chick cochlear nucleus, the nucleus magnocellularis (nMAG), by photolysis of caged glutamate immediately after activation of a single synaptic input. Additionally, synaptic desensitization was demonstrated via competition between synaptically released glutamate and an exogenous nondesensitizing agonist, kainate. Both approaches indicated that at least 35-40% of the receptors were desensitized after a single synaptic stimulus. Miniature synaptic currents were depressed after an evoked synaptic current, indicating that desensitization led to a reduction in the response to individual transmitter quanta. Stimulation of adjacent glutamatergic inputs to the same cell demonstrated that nearby terminals did not depress one another, suggesting that the desensitizing level of glutamate is restricted to each axon terminal. These findings confirm that postsynaptic neurons may use desensitization to regulate the strength of transmission on a synapse-specific basis.
Figures




References
-
- Barbour B, Keller BU, Llano I, Marty A. Prolonged presence of glutamate during excitatory synaptic transmission to cerebellar Purkinje cells. Neuron. 1994;12:1331–1343. - PubMed
-
- Clements J. Transmitter time course in the synaptic cleft: its role in central synaptic function. Trends Neurosci. 1996;19:163–171. - PubMed
-
- Diamond JS, Jahr CE. Asynchronous release of synaptic vesicles determines the time course of the AMPA receptor-mediated EPSC. Neuron. 1995;15:1097–1107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources