

Metaphase spindles rotate in the neuroepithelium of rat cerebral cortex
- PMID: 8922417
- PMCID: PMC6579107
- DOI: 10.1523/JNEUROSCI.16-23-07610.1996
Metaphase spindles rotate in the neuroepithelium of rat cerebral cortex
Abstract
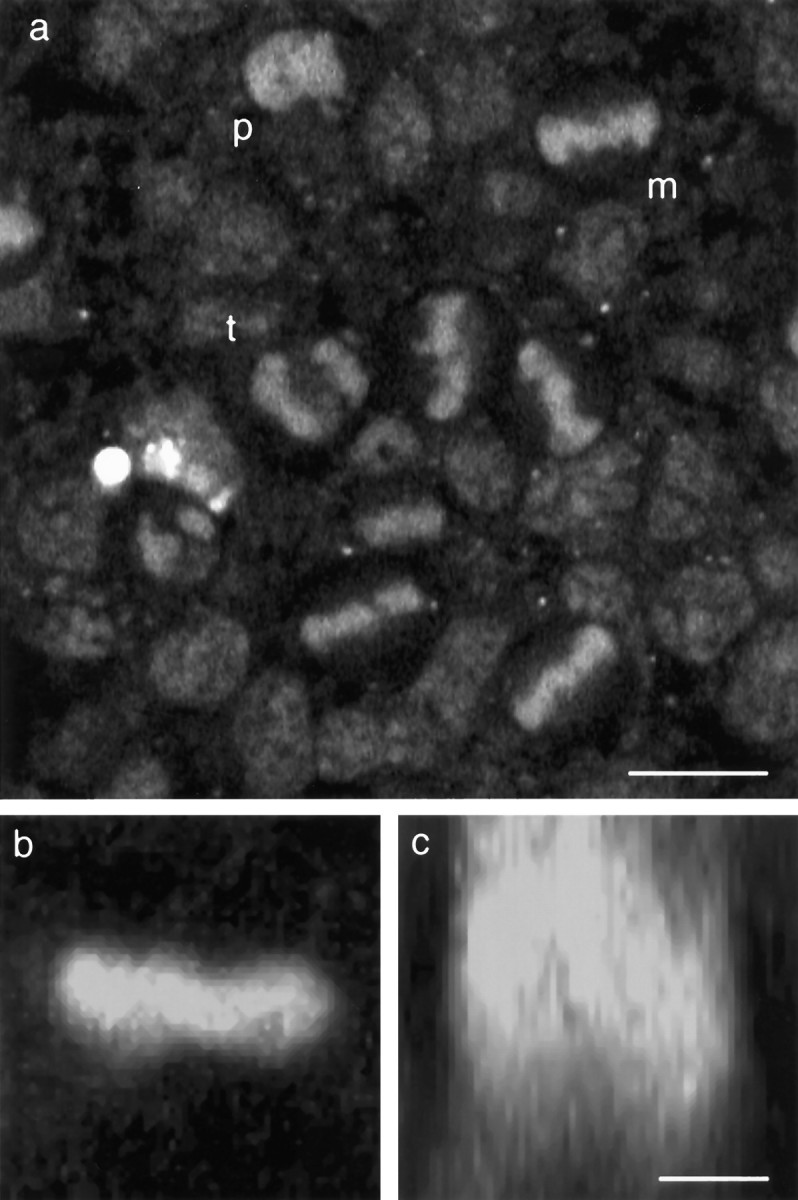
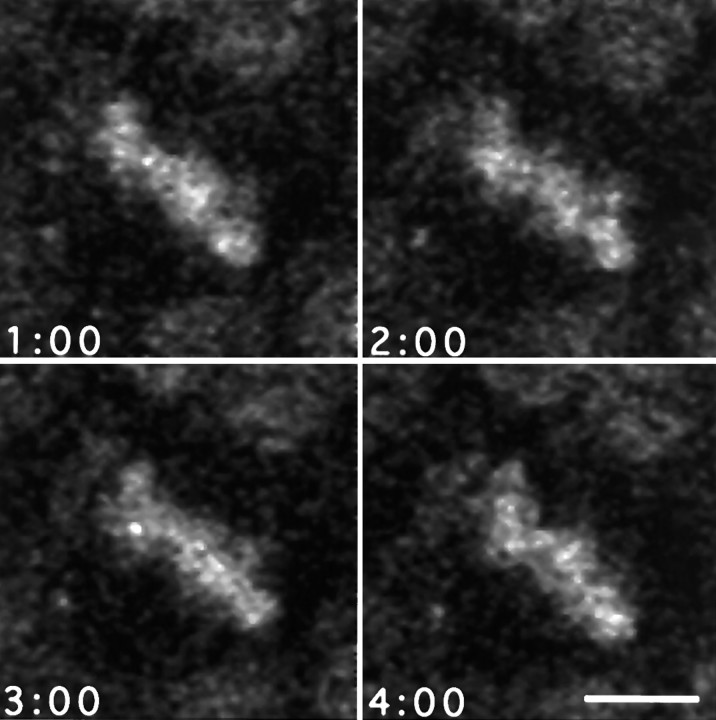
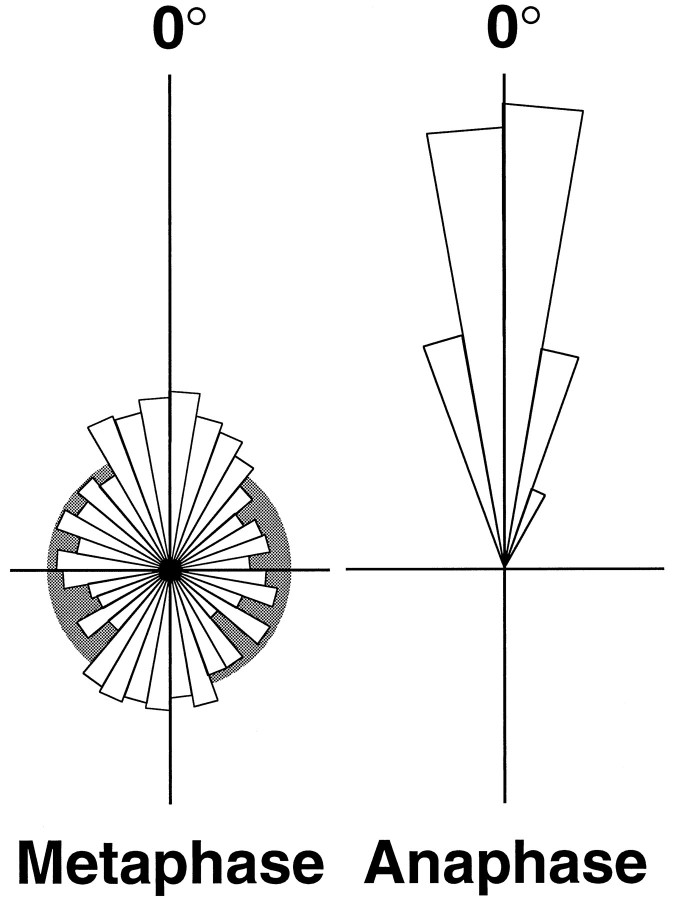
Time-lapse confocal microscopy has been used to image cells in mitosis at the apical surface of neuroepithelium from the rat cerebral cortex during the period of neurogenesis. Staining with vital chromatin dyes reveals that mitotic spindles that are aligned parallel to the surface of the tissue are highly motile, rotating within the plane of the epithelium throughout metaphase, and come to rest only as anaphase begins. Spindles may make several complete turns, parallel to the epithelium, but only rarely tumble into an orientation perpendicular to the epithelial sheet. Analysis shows that spindles do not rotate randomly; rather, they spend most of their time aligned parallel or antiparallel to the direction in which they will later enter anaphase and undergo cell division. This conclusion is strongly supported by statistical analyses of the data. Stereotyped movements of this kind show that the direction of division is determined early in mitosis. This suggests the existence of intracellular and perhaps intercellular signals that define the polarity of the cell both in the apico-basal direction and within the plane of the epithelium. Such mechanisms may be important for maintaining the structure of the epithelium and cell-cell communication during development and may also provide a mechanism for the precise distribution of cytoplasmic determinants that might influence the fate of the daughter cells at a time when neuronal fate is being determined.
Figures


References
-
- Alvarez IS, Martín-Patido G, Rodríguez-Gallardo L, González-Ramos C, Navasués J. Cell proliferation during early development of the chick otic anlage: quantitative comparison of migratory and nonmigratory regions of the otic epithelium. J Comp Neurol. 1989;290:278–288. - PubMed
-
- Barbe MF. Tempting fate and commitment in the developing forebrain. Neuron. 1996;16:1–16. - PubMed
-
- Bayer SA, Altman J. Neocortical development. Raven; New York: 1991.
-
- Chenn A, McConnell SK. Cleavage orientation and the asymmetric inheritance of notch1 immunoreactivity in mammalian neurogenesis. Cell. 1995;82:631–641. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources