

Visual response properties of striate cortical neurons projecting to area MT in macaque monkeys
- PMID: 8922429
- PMCID: PMC6579106
- DOI: 10.1523/JNEUROSCI.16-23-07733.1996
Visual response properties of striate cortical neurons projecting to area MT in macaque monkeys
Abstract
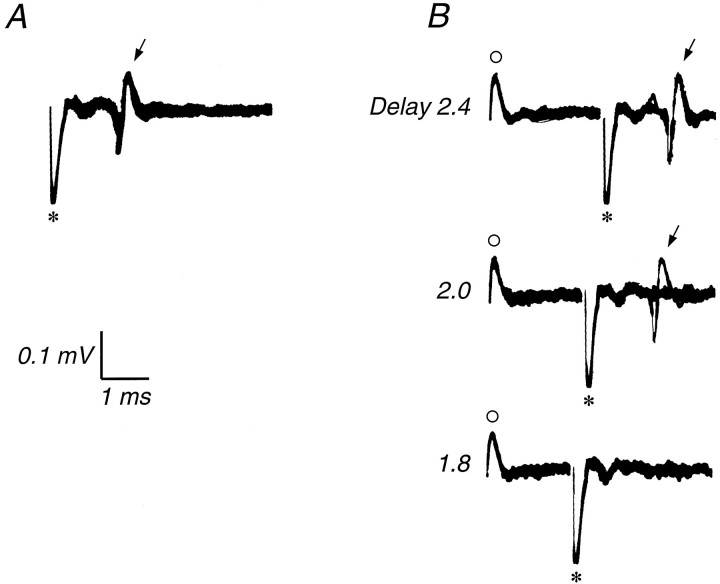
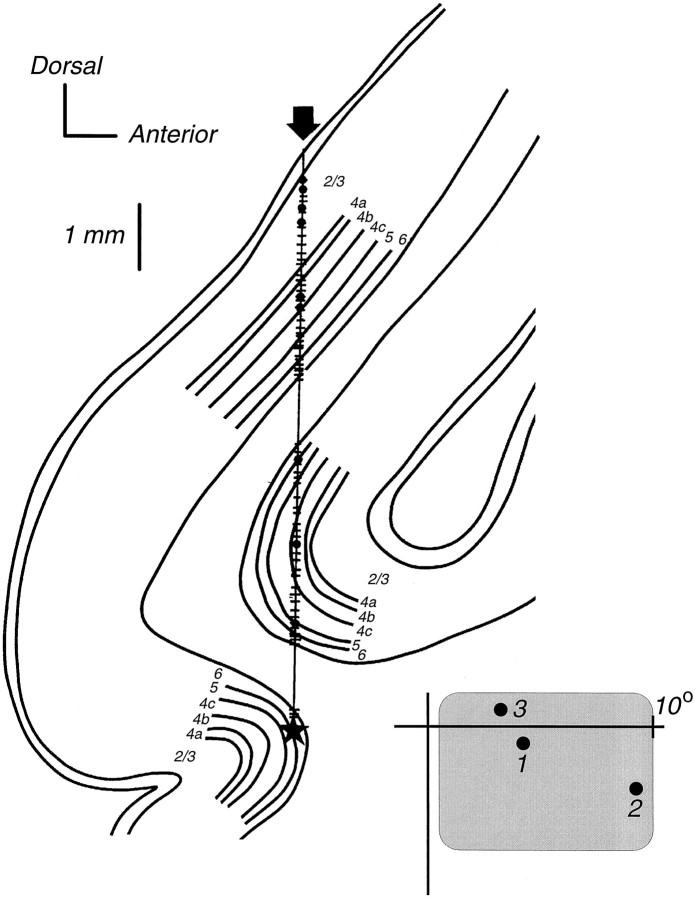
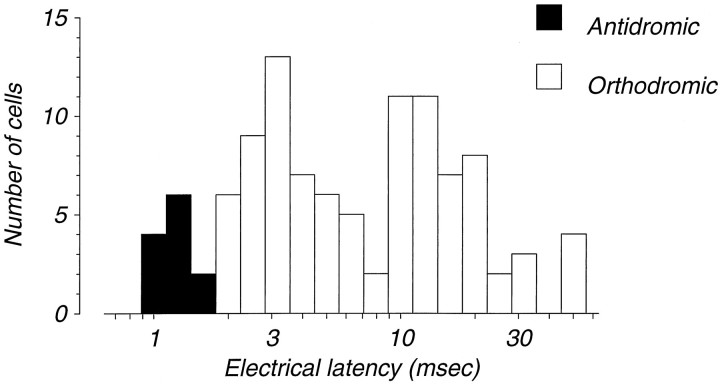
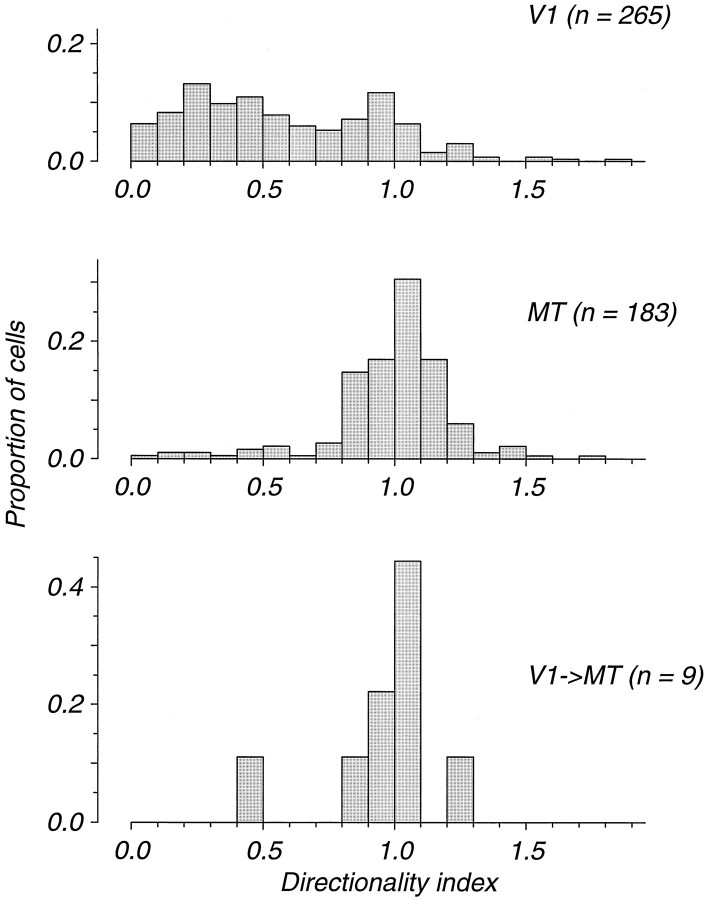
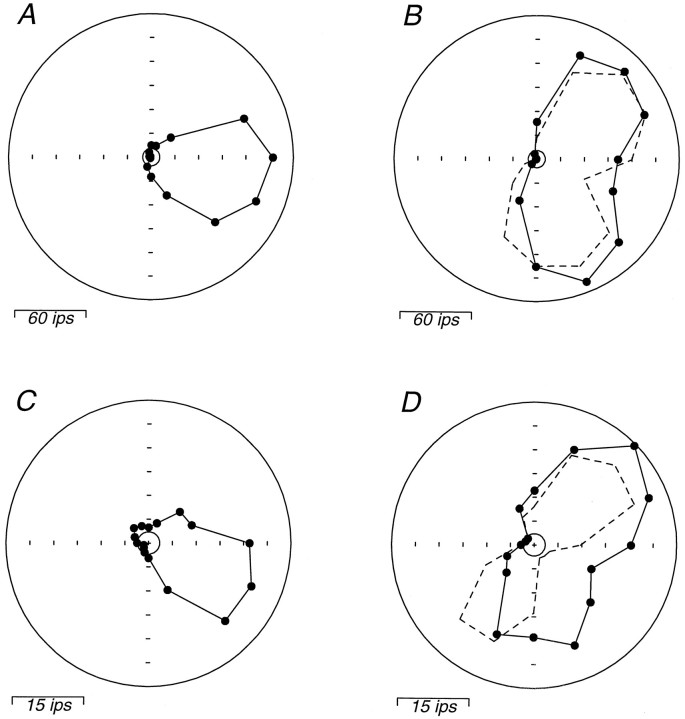
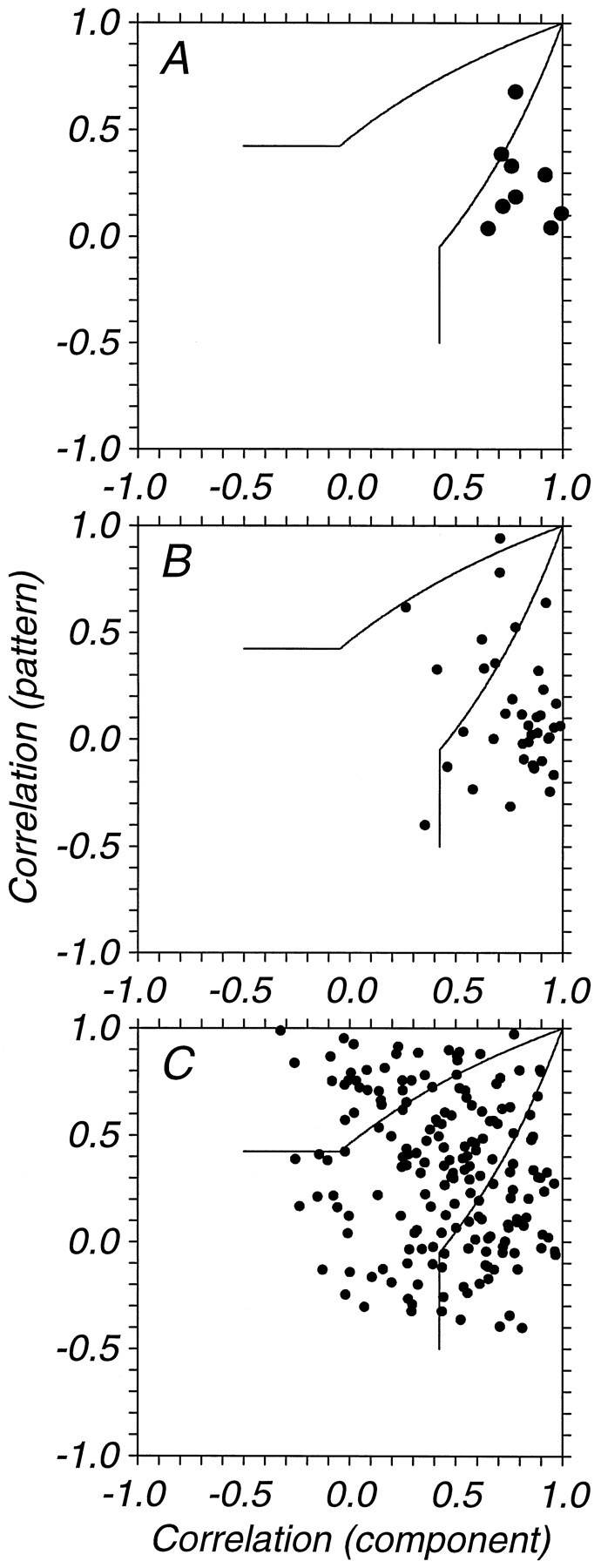
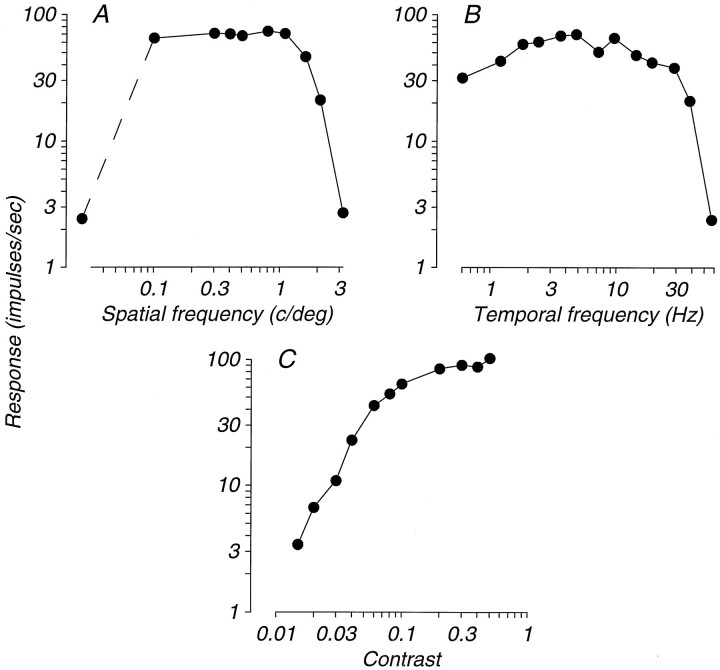
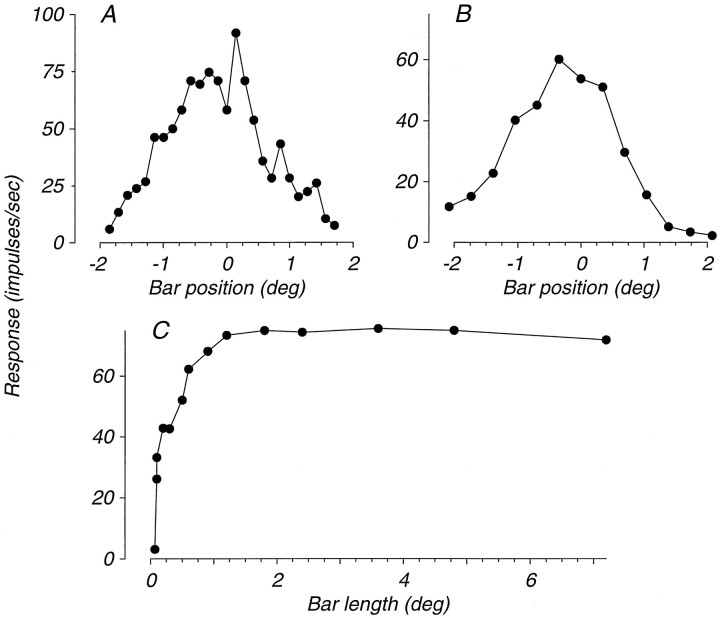
We have previously shown that some neurons in extrastriate area MT are capable of signaling the global motion of complex patterns; neurons randomly sampled from V1, on the other hand, respond only to the motion of individual oriented components. Because only a small fraction of V1 neurons projects to MT, we wished to establish the processing hierarchy more precisely by studying the properties of those neurons projecting to MT, identified by antidromic responses to electrical stimulation of MT. The neurons that project from V1 to MT were directionally selective and, like other V1 neurons, responded only to the motion of the components of complex patterns. The projection neurons were predominantly "special complex," responsive to a broad range of spatial and temporal frequencies, and sensitive to very low stimulus contrasts. The projection neurons thus comprise a homogeneous and highly specialized subset of V1 neurons, consistent with the notion that V1 acts as clearing house of basic visual measurements, distributing information appropriately to higher cortical areas for specialized analysis.
Figures
References
-
- Adelson EH, Bergen JR. Spatio-temporal energy models for the perception of motion. J Opt Soc Am [A] 1985;2:284–299. - PubMed
-
- Adelson EH, Movshon JA. Phenomenal coherence of moving visual patterns. Nature. 1982;300:523–525. - PubMed
-
- Allman JM, Kaas JH. A representation of the visual field in the caudal third of the middle temporal gyrus of the owl monkey (Aotus trivirgatus). Brain Res. 1971;31:85–105. - PubMed
-
- Bender DB. Receptive-field properties of neurons in the macaque inferior pulvinar. J Neurophysiol. 1982;48:1–17. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources