

Molecular determinants of beta 1 subunit-induced gating modulation in voltage-dependent Na+ channels
- PMID: 8929421
- PMCID: PMC6578941
- DOI: 10.1523/JNEUROSCI.16-22-07117.1996
Molecular determinants of beta 1 subunit-induced gating modulation in voltage-dependent Na+ channels
Abstract
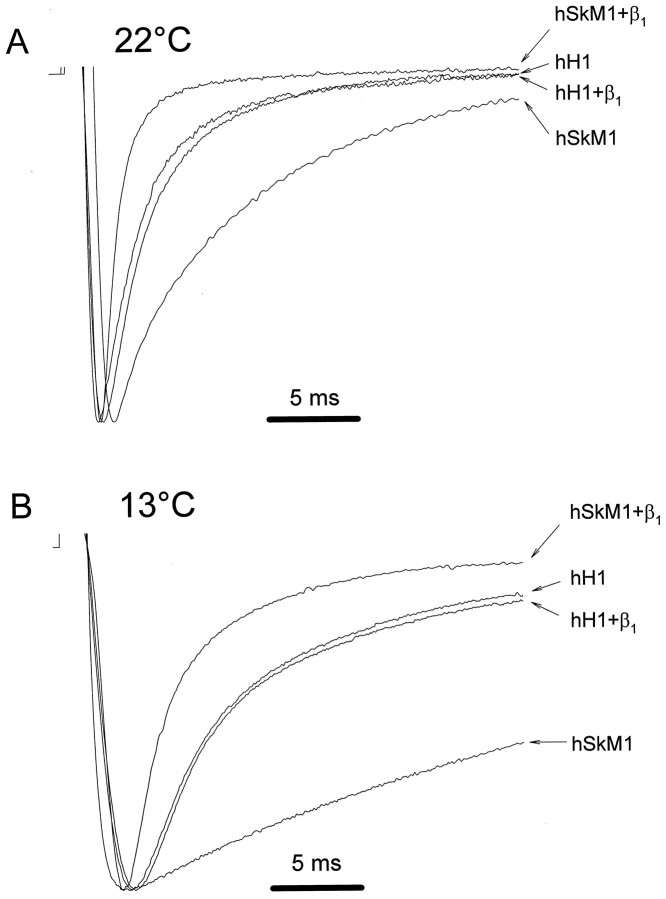
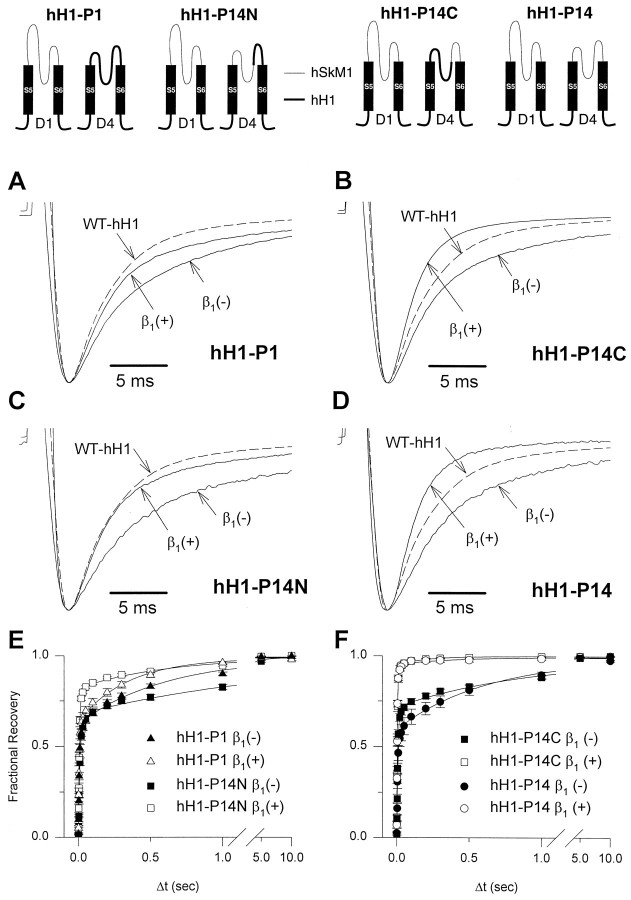
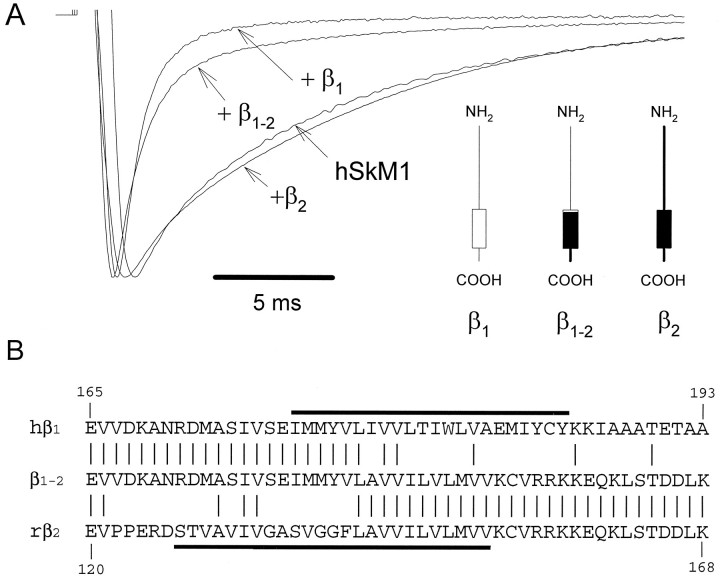
Recombinant brain, skeletal muscle, and heart voltage-gated Na+ channel alpha subunits differ in their functional responses to an accessory beta 1 subunit when coexpressed in Xenopus oocytes. We exploited the distinct beta 1 subunit responses observed for the human heart (hH1) and human skeletal muscle (hSkM1) isoforms to identify determinants of this response. Chimeric alpha subunits were constructed by exchanging the S5-S6 interhelical loops of each domain between hH1 and hSkM1 and then examined for effects on inactivation induced by coexpressed beta 1 subunit in oocytes. Substitution of single S5-S6 loops in either domain 1 (D1/S5-S6) or domain 4 (D4/S5-S6) of hSkM1 by the corresponding segments of hH1 produced channels that exhibited an attenuated response to coexpressed beta 1 subunit. Substitutions of both D1/S5-S6 and D4/S5-S6 in hSkM1 by the corresponding loops from hH1 completely abolished the effects of the beta 1 subunit on inactivation. The reciprocal chimera in which both D1/S5-S6 and D4/S5-S6 from hSkM1 were transplanted into hH1 exhibited significant beta 1 responsiveness (accelerated inactivation). The region within D4/S5-S6 that conferred beta 1 responsiveness was determined to reside primarily within an extracellular loop between the putative pore-forming segment SS2 and D4/S6. Gating modulation was also demonstrated using a chimeric beta subunit consisting of the extracellular domains of beta 1 and the transmembrane and C-terminal domains of the rat brain beta 2 subunit. These results suggest that the D1/S5-S6 and D4/S5-S6 loops in the alpha subunit and the extracellular domain of the beta 1 subunit are important determinants of the beta 1 subunit-induced gating modulation in Na+ channels.
Figures



References
-
- Auld VJ, Goldin AL, Krafte DS, Marshall J, Dunn JM, Catterall WA, Lester HA, Davidson N, Dunn RJ. A rat brain Na+ channel α subunit with novel gating properties. Neuron. 1988;1:449–461. - PubMed
-
- Backx PH, Yue DT, Lawrence JH, Marban E, Tomaselli GF. Molecular localization of an ion-binding site within the pore of mammalian sodium channels. Science. 1992;257:248–251. - PubMed
-
- Bennett PB, Makita N, George AL., Jr A molecular basis for gating mode transitions in human skeletal muscle sodium channels. FEBS Lett. 1993;326:21–24. - PubMed
-
- Cannon SC, McClatchey AI, Gusella JF. Modification of the Na+ current conducted by the rat skeletal muscle α subunit by coexpression with a human brain β subunit. Pflügers Arch. 1993;423:155–157. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources