

Different balance of excitation and inhibition in forward and feedback circuits of rat visual cortex
- PMID: 8929442
- PMCID: PMC6578929
- DOI: 10.1523/JNEUROSCI.16-22-07353.1996
Different balance of excitation and inhibition in forward and feedback circuits of rat visual cortex
Abstract
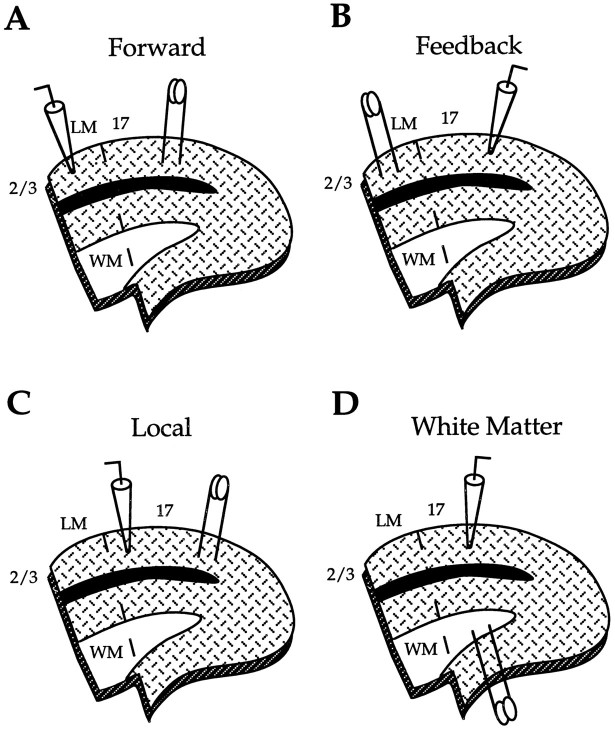
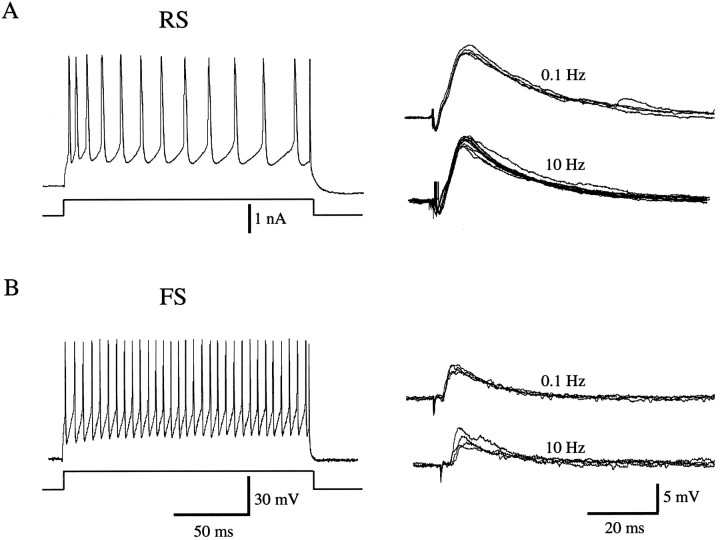
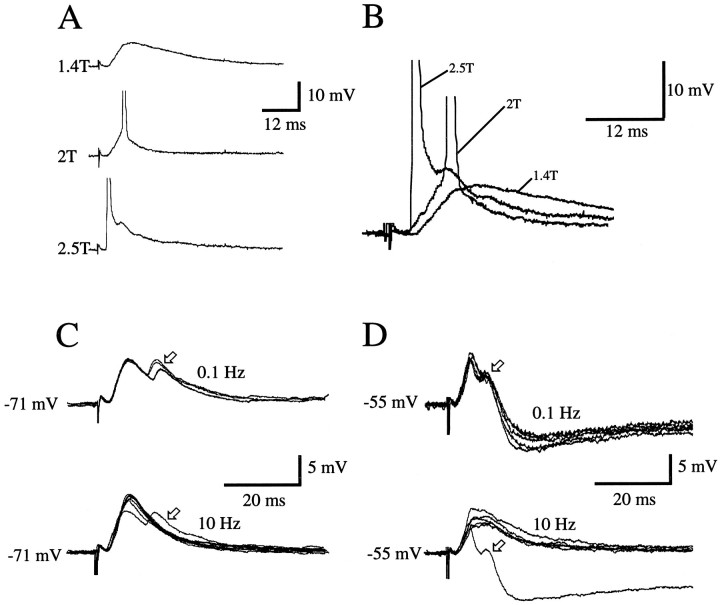
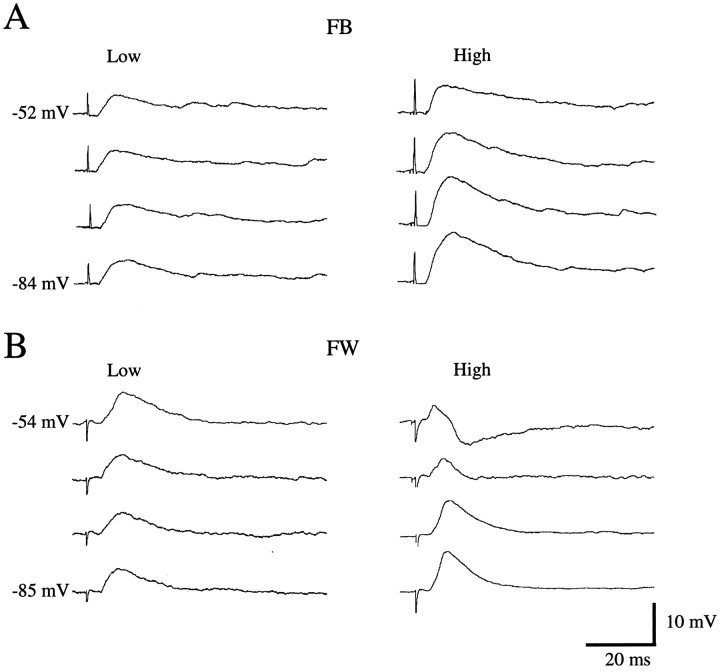
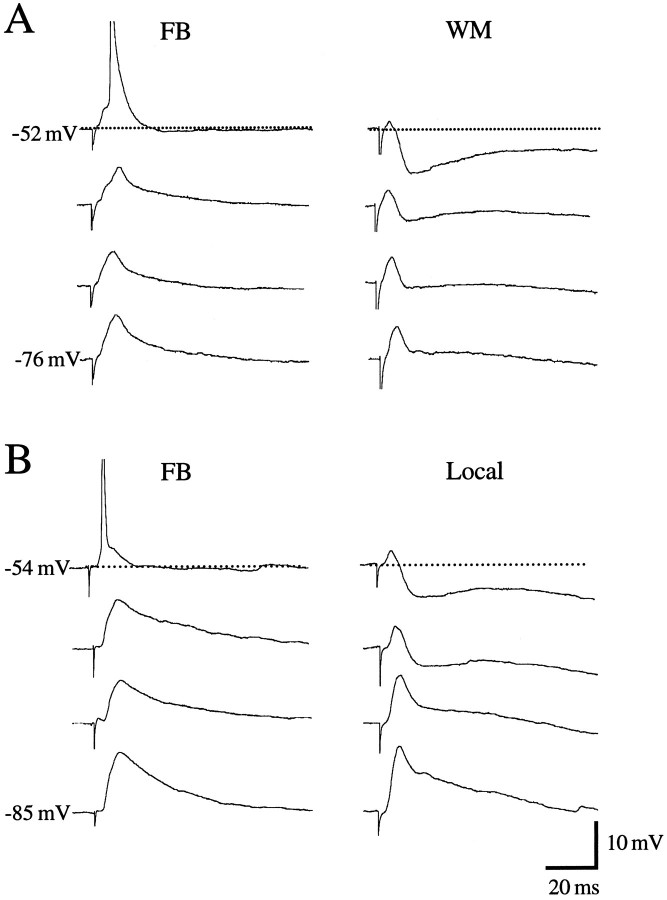
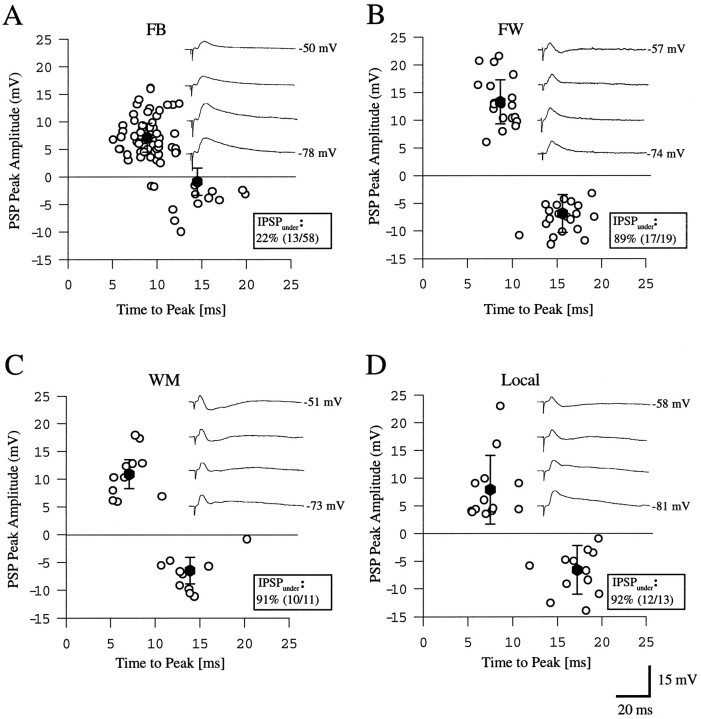
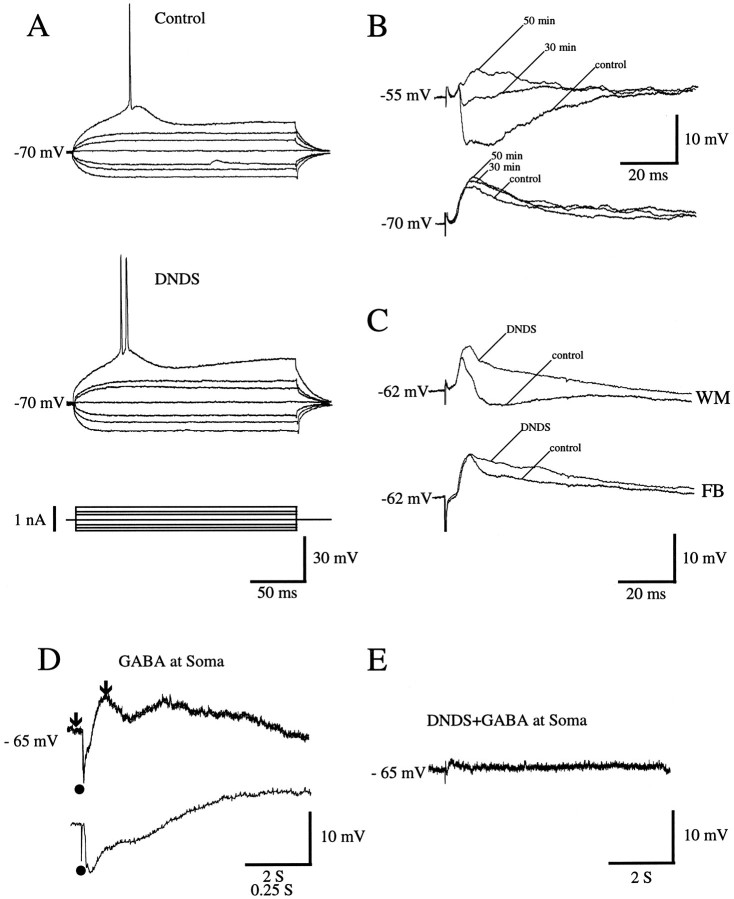
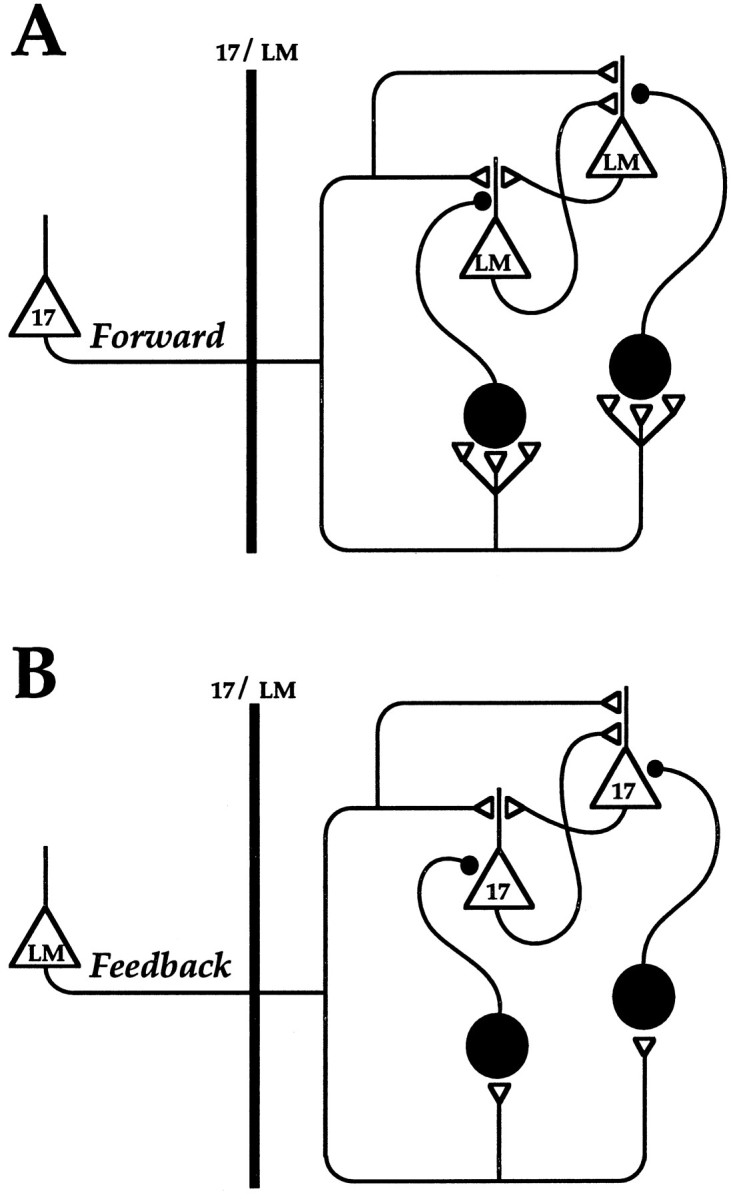
Different cortical areas are linked reciprocally via forward and feedback connections. Forward connections are involved in the representation of retinal images, whereas feedback pathways may play a role in the selection and interpretation of visual information. To examine the synaptic mechanisms of forward and feedback connections between primary and secondary visual cortical areas directly, we have performed intracellular recordings in slices of rat visual cortex. Irrespective of stimulus intensity and membrane potential, 78% (45/58) of the cells in striate cortex activated by feedback input showed monosynaptic responses that were depolarizing only, and inhibitory inputs were evident merely as a slight acceleration in the decay of EPSPs. In contrast, in 89% (17/19) of the cells, stimulation of forward input evoked monosynaptic excitatory postsynaptic potentials (EPSPs), followed by disynaptic, hyperpolarizing inhibitory postsynaptic potentials (IPSPs). EPSPs followed by IPSPs also were recorded after stimulation of local connections within primary visual cortex (92%, 12/13) and after activation of thalamocortical input (91%, 10/11). These results suggest that the synaptic organization of feedback connections are distinct from forward, local, and thalamocortical circuits. The findings further indicate that intracortical back projections exert modulatory influences via synaptic mechanisms in which weak inhibitory input is strongly dominated by excitation.
Figures
References
-
- Aghajanian GK, Rasmussen K. Intracellular studies in the facial nucleus illustrating a simple method for obtaining viable motoneurons in adult brain slices. Synapse. 1989;3:331–338. - PubMed
-
- Allman J, Miezin F, McGuinness E. Stimulus-specific responses from beyond the classical receptive field: neurophysiological mechanisms for local–global comparisons in visual neurons. Annu Rev Neurosci. 1985;8:407–430. - PubMed
-
- Asanuma H, Sakata H. Functional organization of a cortical efferent system examined with focal depth stimulation in cats. J Neurophysiol. 1967;30:35–54.
-
- Buhl EH, Halasy K, Somogyi P. Diverse sources of hippocampal unitary inhibitory postsynaptic potentials and the number of synaptic release sites. Nature. 1994;368:823–828. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources