

Discovery of genes involved with learning and memory: an experimental synthesis of Hirschian and Benzerian perspectives
- PMID: 8942957
- PMCID: PMC33631
- DOI: 10.1073/pnas.93.24.13460
Discovery of genes involved with learning and memory: an experimental synthesis of Hirschian and Benzerian perspectives
Abstract
The biological bases of learning and memory are being revealed today with a wide array of molecular approaches, most of which entail the analysis of dysfunction produced by gene disruptions. This perspective derives both from early "genetic dissections" of learning in mutant Drosophila by Seymour Benzer and colleagues and from earlier behavior-genetic analyses of learning and in Diptera by Jerry Hirsh and coworkers. Three quantitative-genetic insights derived from these latter studies serve as guiding principles for the former. First, interacting polygenes underlie complex traits. Consequently, learning/memory defects associated with single-gene mutants can be quantified accurately only in equilibrated, heterogeneous genetic backgrounds. Second, complex behavioral responses will be composed of genetically distinct functional components. Thus, genetic dissection of complex traits into specific biobehavioral properties is likely. Finally, disruptions of genes involved with learning/memory are likely to have pleiotropic effects. As a result, task-relevant sensorimotor responses required for normal learning must be assessed carefully to interpret performance in learning/memory experiments. In addition, more specific conclusions will be obtained from reverse-genetic experiments, in which gene disruptions are restricted in time and/or space.
Figures
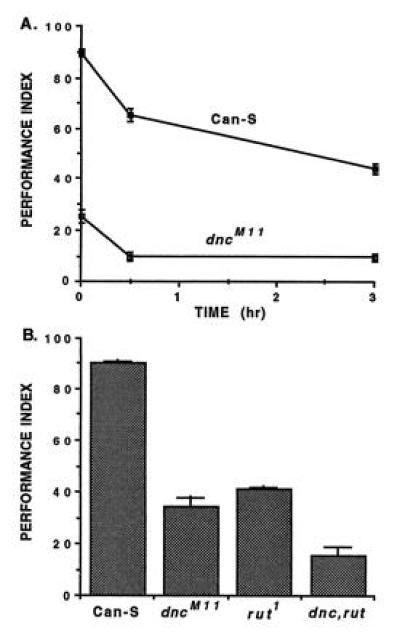
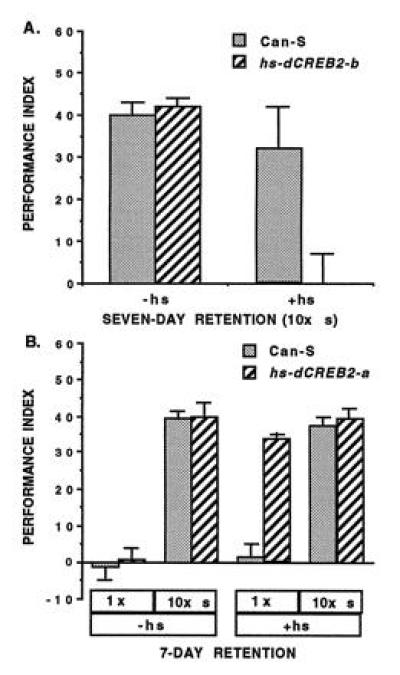
References
-
- Darwin C. The Descent of Man and Selection in Relation to Sex. London: John Murray; 1871.
-
- Galton F. Hereditary Genius: An Inquiry into Its Laws and Consequences. London: Macmillan; 1869.
-
- Hall C S. In: Handbook of Experimental Psychology. Stevens S S, editor. New York: Wiley; 1951. pp. 304–329.
-
- Tolman E C. J Comp Psychol. 1924;4:1–18.
-
- Tryon R C. Yearbook Natl Soc Stud Edu. 1940;39:111–119.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
