

Involvement of the cyclin-dependent kinase inhibitor p16 (INK4a) in replicative senescence of normal human fibroblasts
- PMID: 8943005
- PMCID: PMC19411
- DOI: 10.1073/pnas.93.24.13742
Involvement of the cyclin-dependent kinase inhibitor p16 (INK4a) in replicative senescence of normal human fibroblasts
Abstract
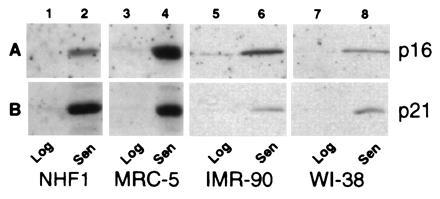
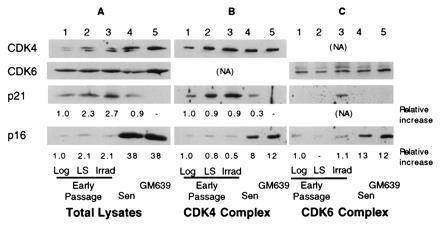
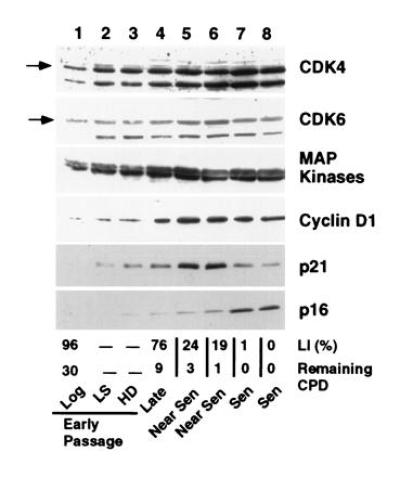
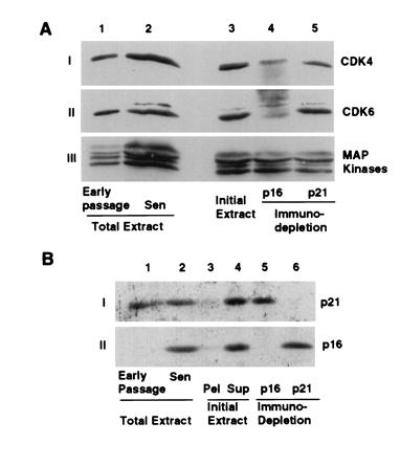
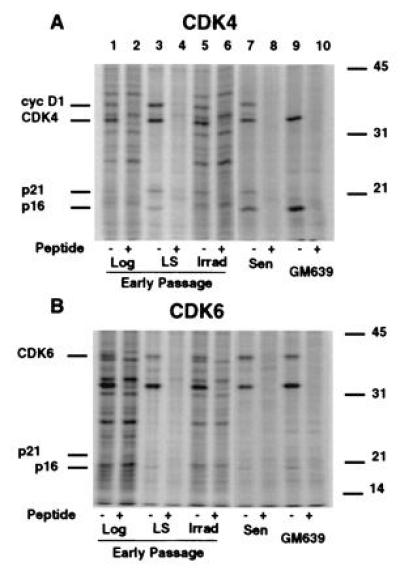
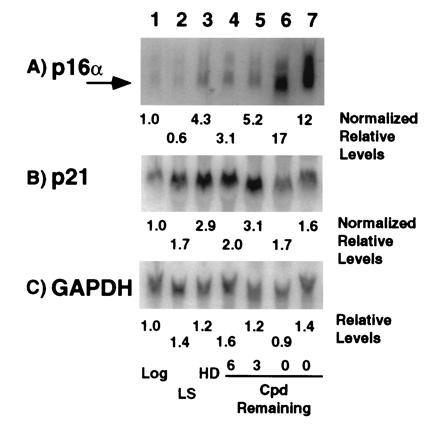
Human diploid fibroblasts (HDFs) can be grown in culture for a finite number of population doublings before they cease proliferation and enter a growth-arrest state termed replicative senescence. The retinoblastoma gene product, Rb, expressed in these cells is hypophosphorylated. To determine a possible mechanism by which senescent human fibroblasts maintain a hypophosphorylated Rb, we examined the expression levels and interaction of the Rb kinases, CDK4 and CDK6, and the cyclin-dependent kinase inhibitors p21 and p16 in senescent HDFs. Cellular p21 protein expression increased dramatically during the final two to three passages when the majority of cells lost their growth potential and neared senescence but p21 levels declined in senescent HDFs. During this period, p16 mRNA and cellular protein levels gradually rose with the protein levels in senescent HDFs reaching nearly 40-fold higher than early passage cells. In senescent HDFs, p16 was shown to be complexed to both CDK4 and CDK6. Immunodepletion analysis of p21 and p16 from the senescent cell extracts revealed that p16 is the major CDK inhibitor for both CDK4 and CDK6 kinases. Immunoprecipitation of CDK4 and CDK6 and their associated proteins from radiolabeled extracts from senescent HDFs showed no other CDK inhibitors. Based upon these results, we propose that senescence is a multistep process requiring the expression of both p21 and p16. p16 up-regulation is a key event in the terminal stages of growth arrest in senescence, which may explain why p16 but not p21 is commonly mutated in immortal cells and human tumors.
Figures
References
-
- Hayflick L. Exp Cell Res. 1965;37:614–636. - PubMed
-
- Shay J W, Pereira-Smith O M, Wright W E. Exp Cell Res. 1991;196:33–39. - PubMed
-
- Hara E, Tsurui H, Shinozaki A, Nakada S, Oda K. Biochem Biophys Res Commun. 1991;179:528–534. - PubMed
-
- Stein G H, Beeson M, Gordon L. Science. 1990;249:666–669. - PubMed
-
- Weinberg R A. Cell. 1995;81:323–330. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
