

Molecular characterization of the human peroxisomal branched-chain acyl-CoA oxidase: cDNA cloning, chromosomal assignment, tissue distribution, and evidence for the absence of the protein in Zellweger syndrome
- PMID: 8943006
- PMCID: PMC19413
- DOI: 10.1073/pnas.93.24.13748
Molecular characterization of the human peroxisomal branched-chain acyl-CoA oxidase: cDNA cloning, chromosomal assignment, tissue distribution, and evidence for the absence of the protein in Zellweger syndrome
Abstract
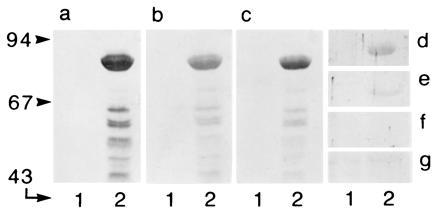
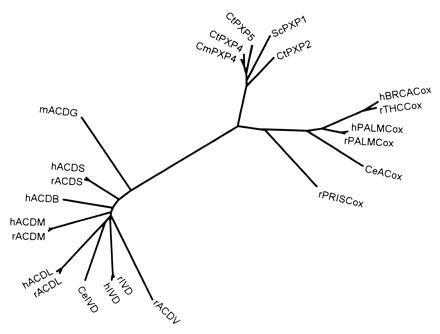
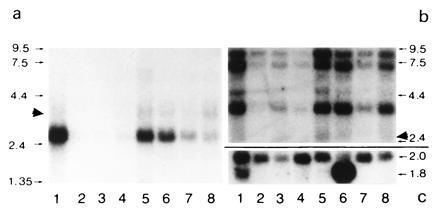
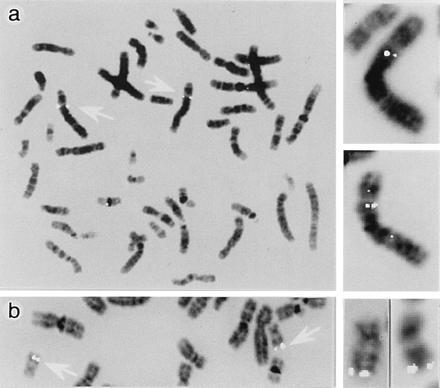
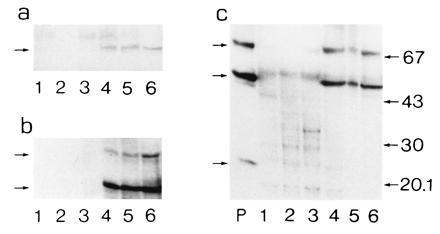
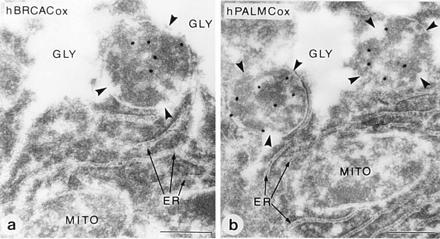
Peroxisomes in human liver contain two distinct acyl-CoA oxidases with different substrate specificities: (i) palmitoyl-CoA oxidase, oxidizing very long straight-chain fatty acids and eicosanoids, and (ii) a branched-chain acyl-CoA oxidase (hBRCACox), involved in the degradation of long branched fatty acids and bile acid intermediates. The accumulation of branched fatty acids and bile acid intermediates leads to severe mental retardation and death of the diseased children. In this study, we report the molecular characterization of the hBRCACox, a prerequisite for studying mutations in patients with a single enzyme deficiency. The composite cDNA sequence of hBRCACox, derived from overlapping clones isolated via immunoscreening and hybridization of human liver cDNA expression libraries, consisted of 2225 bases and contained an open reading frame of 2046 bases, encoding a protein of 681 amino acids with a calculated molecular mass of 76,739 Da. The C-terminal tripeptide of the protein is SKL, a known peroxisome targeting signal. Sequence comparison with the other acyl-CoA oxidases and evolutionary analysis revealed that, despite its broader substrate specificity, the hBRCACox is the human homolog of rat trihydroxycoprostanoyl-CoA oxidase (rTHCCox) and that separate gene duplication events led to the occurrence in mammals of acyl-CoA oxidases with different substrate specificities. Northern blot analysis demonstrated that--in contrast to the rTHCCox gene--the hBRCACox gene is transcribed also in extrahepatic tissues such as heart, kidney, skeletal muscle, and pancreas. The highest levels of the 2.6-kb mRNA were found in heart, followed by liver. The enzyme is encoded by a single-copy gene, which was assigned to chromosome 3p14.3 by fluorescent in situ hybridization. It was absent from livers of Zellweger patients as shown by immunoblot analysis and immunocytochemistry.
Figures

References
-
- Mannaerts, G. P. & Van Veldhoven, P. P. (1995) Ann. N.Y. Acad. Sci. 804, in press. - PubMed
-
- van den Bosch H, Schutgens R B H, Wanders R J A, Tager J. Annu Rev Biochem. 1992;61:157–197. - PubMed
-
- Reddy J K, Mannaerts G P. Annu Rev Nutr. 1994;14:343–370. - PubMed
-
- Casteels M, Schepers L, Van Veldhoven P P, Eyssen H J, Mannaerts G P. J Lipid Res. 1990;31:1865–1872. - PubMed
-
- Vanhove G F, Van Veldhoven P P, Fransen M, Denis S, Eyssen H J, Wanders R J A, Mannaerts G P. J Biol Chem. 1993;268:10335–10344. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
