

Tumor necrosis factor receptor-associated factor (TRAF)-1, TRAF-2, and TRAF-3 interact in vivo with the CD30 cytoplasmic domain; TRAF-2 mediates CD30-induced nuclear factor kappa B activation
- PMID: 8943059
- PMCID: PMC19493
- DOI: 10.1073/pnas.93.24.14053
Tumor necrosis factor receptor-associated factor (TRAF)-1, TRAF-2, and TRAF-3 interact in vivo with the CD30 cytoplasmic domain; TRAF-2 mediates CD30-induced nuclear factor kappa B activation
Retraction in
-
Tumor necrosis factor receptor-associated factor (TRAF)-1, TRAF-2, and TRAF-3 interact in vivo with the CD30 cytoplasmic domain; TRAF-2 mediates CD30-induced nuclear factor kappa B activation.Proc Natl Acad Sci U S A. 1997 Nov 11;94(23):12732. Proc Natl Acad Sci U S A. 1997. PMID: 9411448 Free PMC article. No abstract available.
Abstract
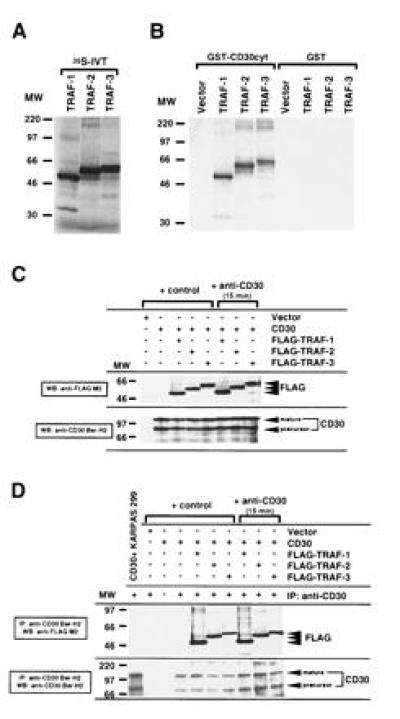
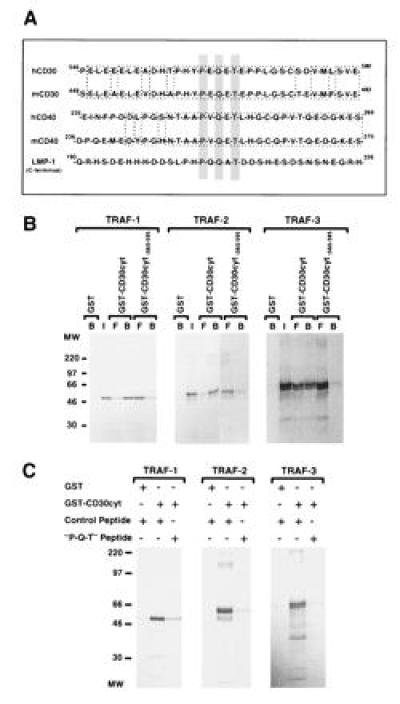
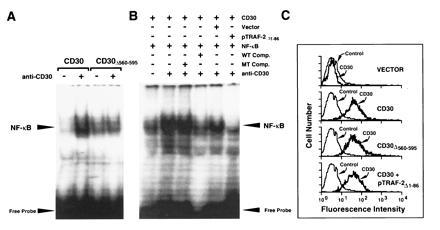
CD30 is a member of the tumor necrosis factor receptor superfamily, which can transduce signals for proliferation, death, or nuclear factor kappa B (NF-kappa B) activation. Investigation of CD30 signaling pathways using a yeast two-hybrid interaction system trapped a cDNA encoding the tumor necrosis factor receptor-associated factor (TRAF)-2 TRAF homology domain. TRAF-1 and TRAF-3 also interacted with CD30, and > 90% of in vitro-translated TRAF-1 or -2, or 50% of TRAF-3, bound to the CD30 cytoplasmic domain. TRAF-1, -2, and -3 bound mostly, but not exclusively, to the carboxyl-terminal 36 residues of CD30. The binding was strongly inhibited by a CD30 oligopeptide centered around a PXQXT (where X is any amino acid) motif shared with CD40 and the Epstein-Barr virus transforming protein LMP1, indicating that this motif in CD30 is an important determinant of TRAF-1, -2 or -3 interaction. At least 15% of TRAF-1, -2, or -3 associated with CD30 when coexpressed in 293 cells. The association was not affected by CD30 cross-linking. However, cross-linking of CD30 activated NF-kappa B. NF-kappa B activation was dependent on the carboxyl-terminal 36 amino acids of CD30 that mediate TRAF association. TRAF-2 has been previously shown to have a unique role in TRAF-mediated NF-kappa B activation, and NF-kappa B activation following CD30 cross-linking was blocked by a dominant negative TRAF-2 mutant. These data indicate that CD30 cross-linking-induced NF-kappa B activation is predominantly TRAF-2-mediated.
Figures
References
-
- Dürkop H, Latza U, Hummel M, Eitelbach F, Seed B, Stein H. Cell. 1992;68:421–427. - PubMed
-
- Smith C A, Farrah T, Goodwin R G. Cell. 1994;75:959–962. - PubMed
-
- Falini B, Pileri S, Pizzolo G, Dürkop H, Flenghi L, Stirpe F, Martelli M F, Stein H. Blood. 1995;85:1–14. - PubMed
-
- Schwab U, Stein H, Gerdes J, Lemke H, Kirchner H, Schaadt M, Diehl V. Nature (London) 1982;299:65–67. - PubMed
-
- Ellis T M, Simms P E, Slivnick D J, Jäck H-M, Fisher R I. J Immunol. 1993;151:2380–2389. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
