

A small nucleolar RNA requirement for site-specific ribose methylation of rRNA in Xenopus
- PMID: 8962077
- PMCID: PMC26158
- DOI: 10.1073/pnas.93.25.14480
A small nucleolar RNA requirement for site-specific ribose methylation of rRNA in Xenopus
Abstract
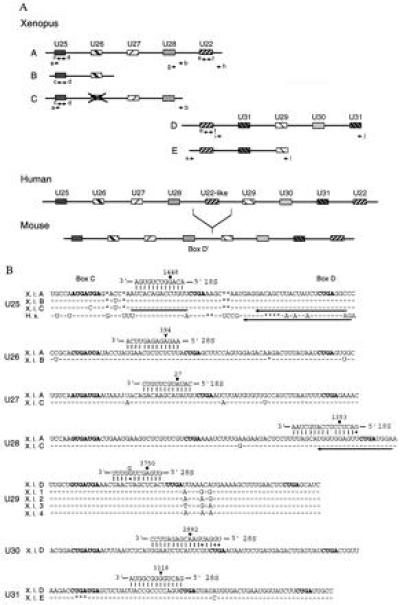
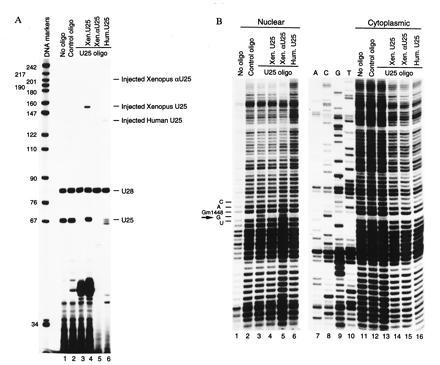
Vertebrate cells contain a large number of small nucleolar RNA (snoRNA) species, the vast majority of which bind fibrillarin. Most of the fibrillarin-associated snoRNAs can form 10- to 21-nt duplexes with rRNA and are thought to guide 2'-O-methylation of selected nucleotides in rRNA. These include mammalian UHG (U22 host gene)-encoded U25-U31 snoRNAs. We have characterized two novel human snoRNA species, U62 and U63, which similarly exhibit 15- (with one interruption) and 12-nt complementarities and are therefore predicted to direct 2'-O-methylation of A590 in 18S and A4531 in 28S rRNA, respectively. To establish the function of antisense snoRNAs in vertebrates, we exploited the Xenopus oocyte system. Cloning of the Xenopus U25-U31 snoRNA genes indicated that they are encoded within multiple homologs of mammalian UHG. Depletion of U25 from the Xenopus oocyte abolished 2'-O-methylation of G1448 in 18S rRNA; methylation could be restored by injecting either the Xenopus or human U25 transcript into U25-depleted oocytes. Comparison of Xenopus and human U25 sequences revealed that only boxes C, D, and D', as well as the 18S rRNA complement, were invariant, suggesting that they may be the only elements required for U25 snoRNA stability and function.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
