

Lineage commitment in the thymus: only the most differentiated (TCRhibcl-2hi) subset of CD4+CD8+ thymocytes has selectively terminated CD4 or CD8 synthesis
- PMID: 8976166
- PMCID: PMC2196385
- DOI: 10.1084/jem.184.6.2091
Lineage commitment in the thymus: only the most differentiated (TCRhibcl-2hi) subset of CD4+CD8+ thymocytes has selectively terminated CD4 or CD8 synthesis
Abstract
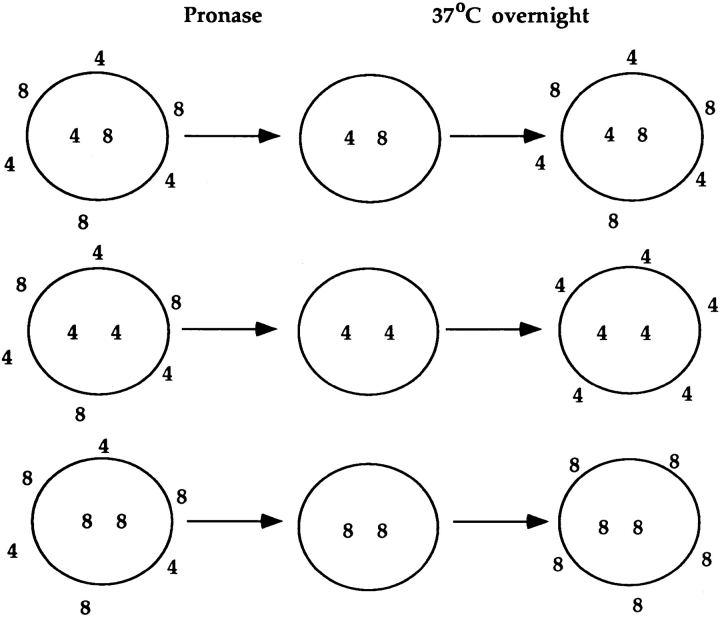
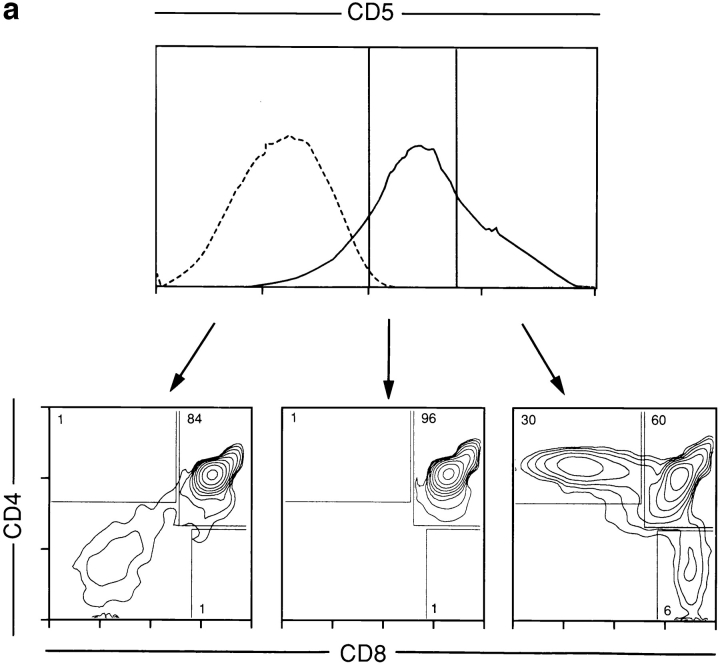
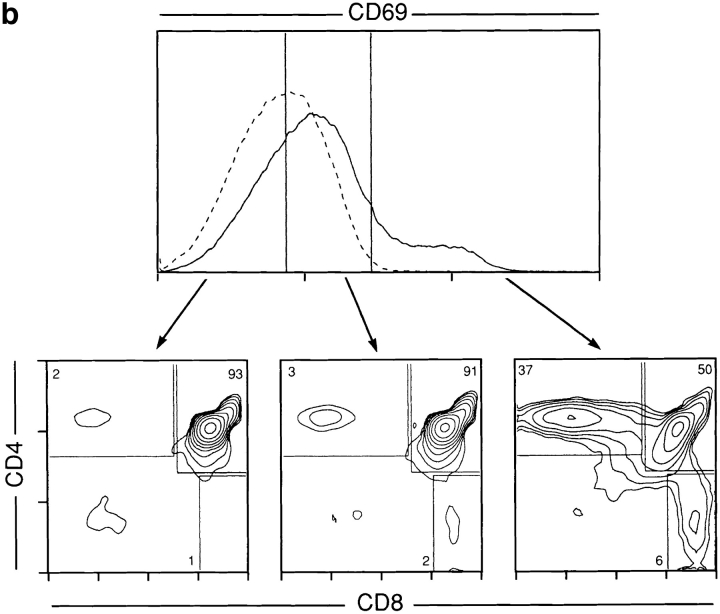
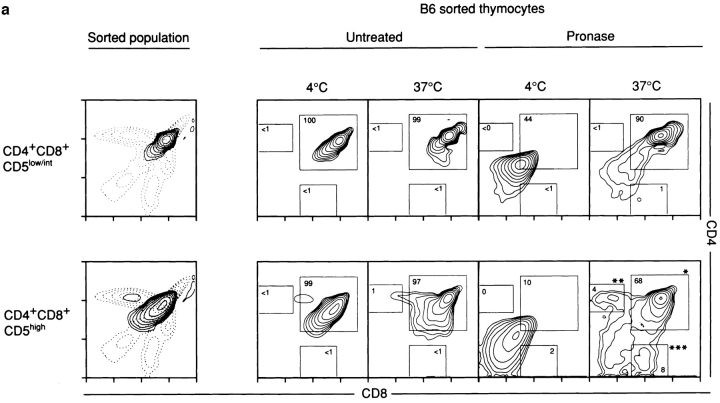
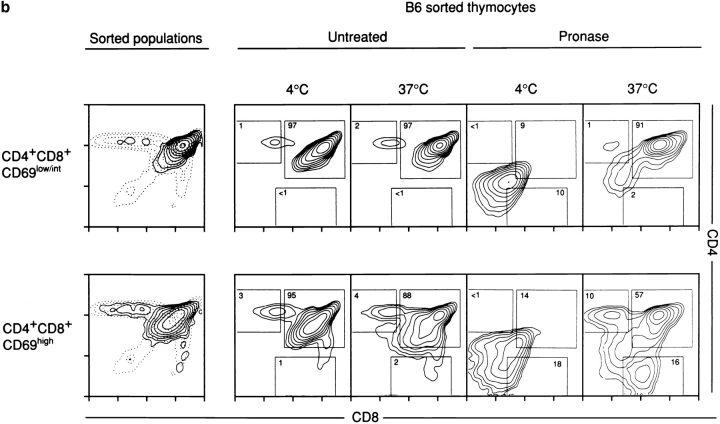
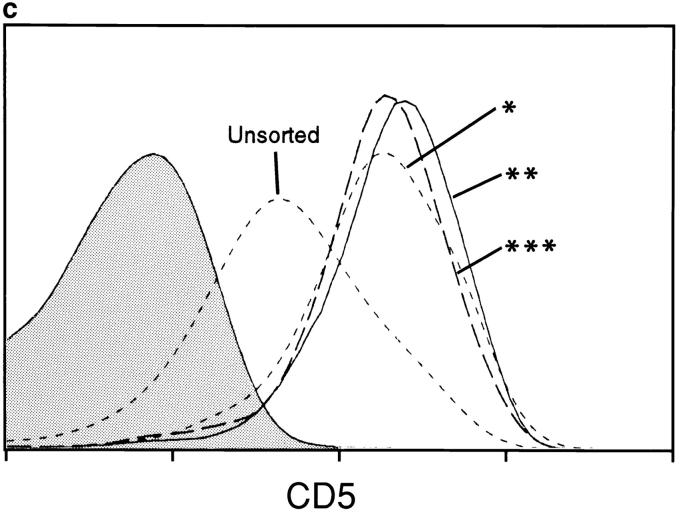
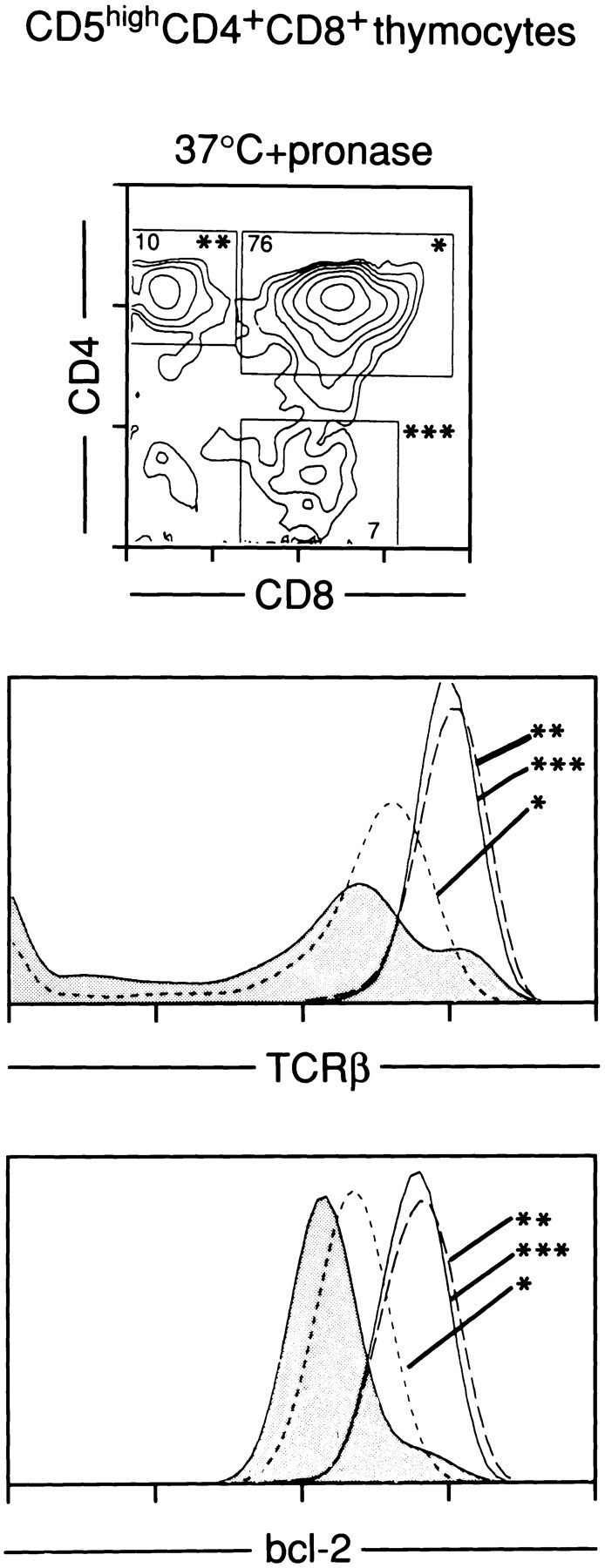
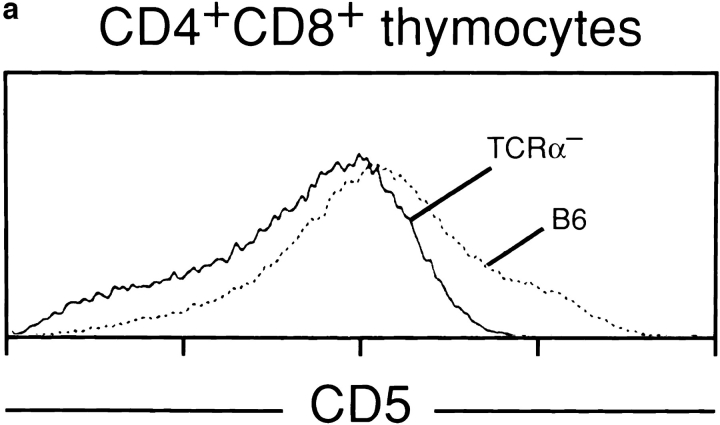
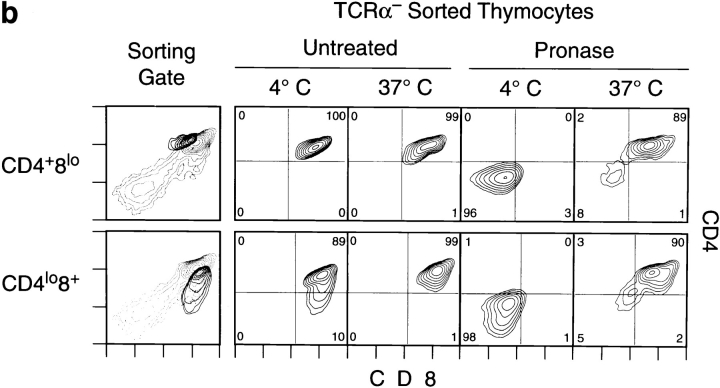
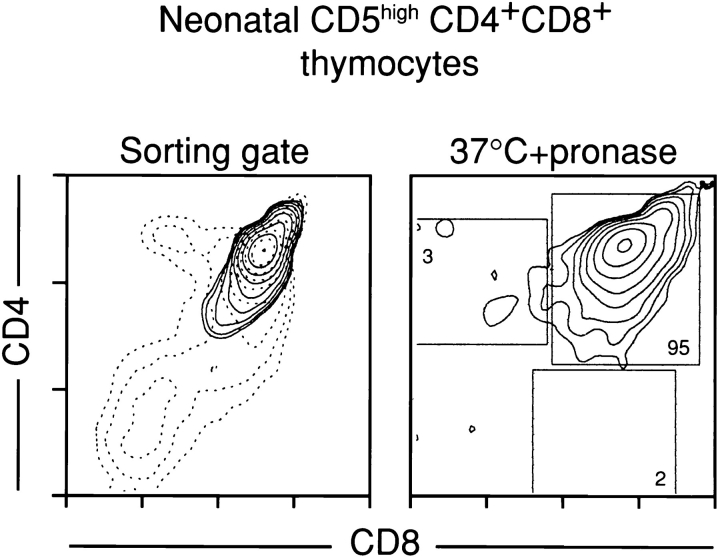
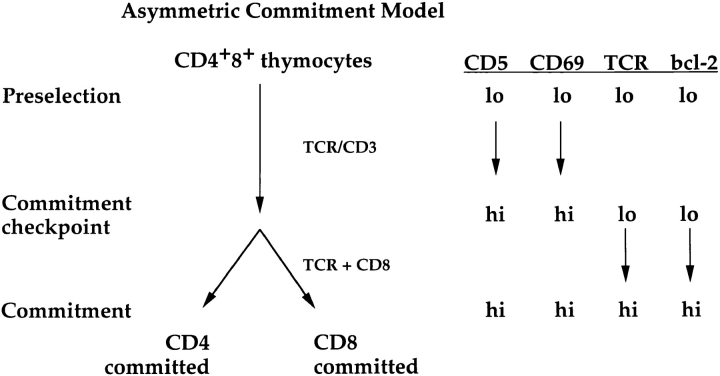
Lineage commitment is a developmental process by which individual CD4+CD8+ (double positive, DP) thymocytes make a decision to differentiate into either CD4+ or CD8+ T cells. However, the molecular event(s) that defines lineage commitment is controversial. We have previously proposed that lineage commitment in DP thymocytes can be molecularly defined as the selective termination of CD4 or CD8 coreceptor synthesis. The present study supports such a molecular definition by showing that termination of either CD4 or CD8 synthesis is a highly regulated event that is only evident within the most differentiated DP subset (CD5hiCD69hiTCRhibcl-2hi). In fact, essentially all cells within this DP subset actively synthesize only one coreceptor molecule. In addition, the present results identify three distinct sub-populations of DP thymocytes that define the developmental progression of the lineage commitment process and demonstrate that lineage commitment is coincident with upregulation of TCR and bcl-2. Thus, this study supports a molecular definition of lineage commitment and uniquely identifies TCRhibcl-2hi DP thymocytes as cells that are already committed to either the CD4 or CD8 T cell lineage.
Figures
References
-
- Robey E, Fowlkes BJ. Selective events in T cell development. Annu Rev Immunol. 1994;12:675–705. - PubMed
-
- Fowlkes BJ, Schweighoffer E. Positive selection of T cells. Curr Opin Immunol. 1995;5:873–879. - PubMed
-
- Davis CB, Killeen N, Crooks ME, Raulet D, Littman DR. Evidence for a stochastic mechanism in the differentiation of mature subsets of T lymphocytes. Cell. 1993;73:237–247. - PubMed
-
- Chan SH, Cosgrove D, Waltzinger C, Benoist C, Mathis D. Another view of the selective model of thymocyte selection. Cell. 1993;73:225–236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
