

HLA-DM interactions with intermediates in HLA-DR maturation and a role for HLA-DM in stabilizing empty HLA-DR molecules
- PMID: 8976171
- PMCID: PMC2196380
- DOI: 10.1084/jem.184.6.2153
HLA-DM interactions with intermediates in HLA-DR maturation and a role for HLA-DM in stabilizing empty HLA-DR molecules
Abstract
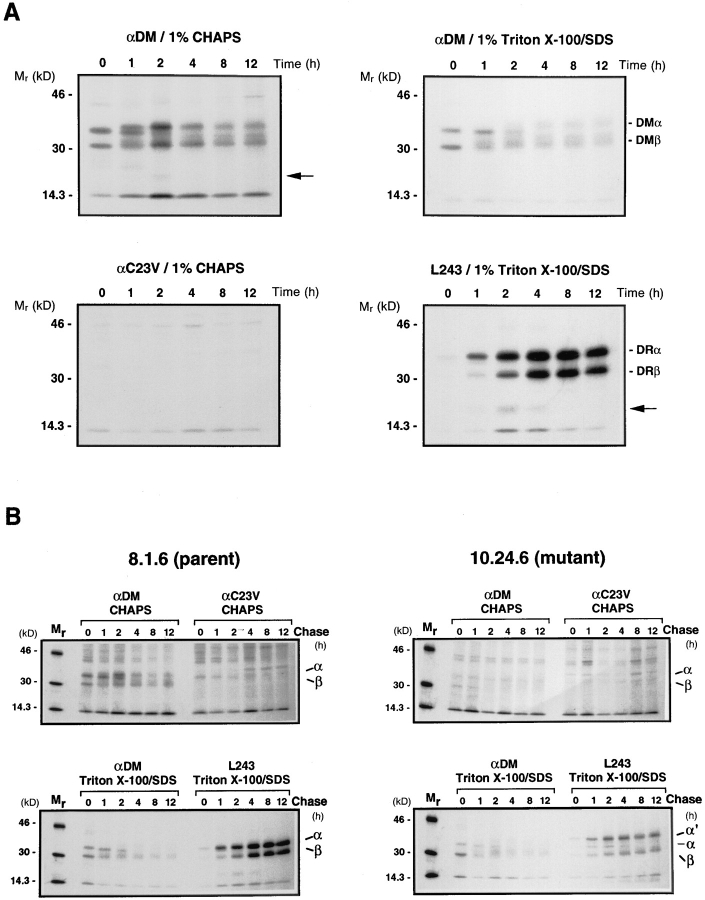
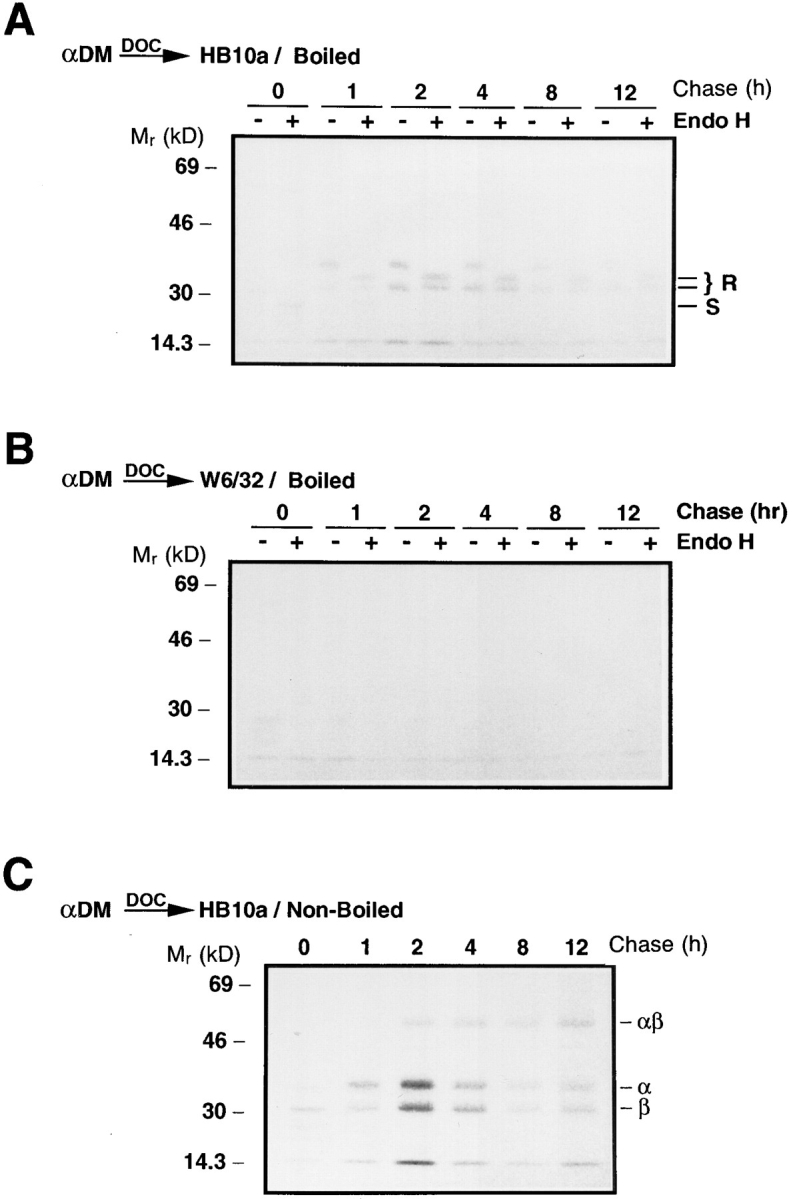
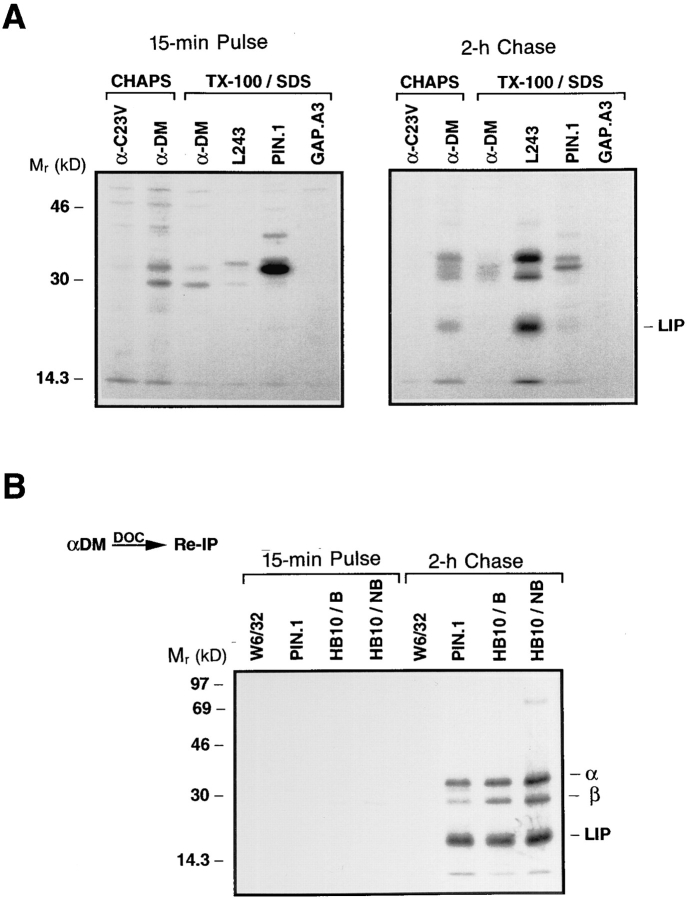
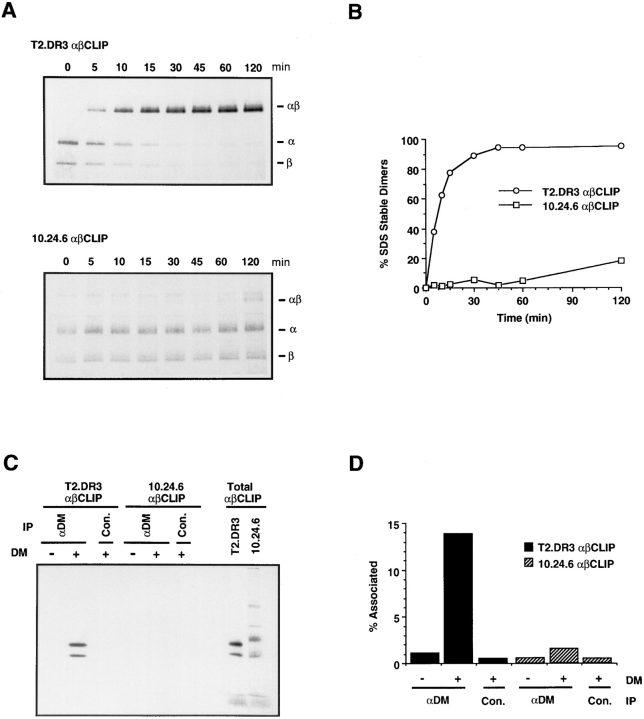
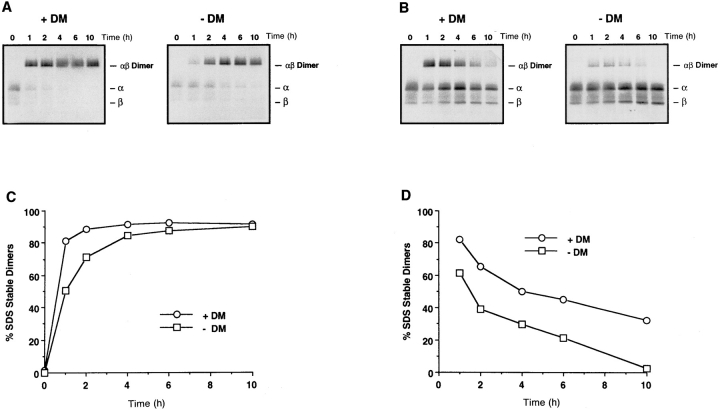
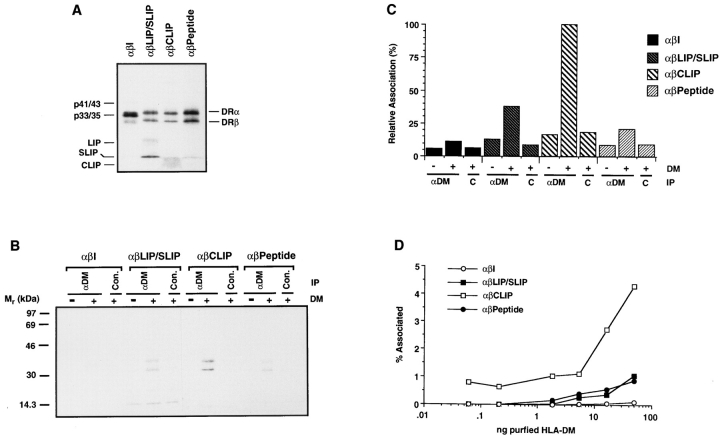
Major histocompatibility complex (MHC) class II-positive cell lines which lack HLA-DM expression accumulate class II molecules associated with residual invariant (I) chain fragments (class II-associated invariant chain peptides [CLIP]). In vitro, HLA-DM catalyzes CLIP dissociation from class II-CLIP complexes, promoting binding of antigenic peptides. Here the physical interaction of HLA-DM with HLA-DR molecules was investigated. HLA-DM complexes with class II molecules were detectable transiently in cells, peaking at the time when the class II molecules entered the MHC class II compartment. HLA-DR alpha beta dimers newly released from I chain, and those associated with I chain fragments, were found to associate with HLA-DM in vivo. Mature, peptide-loaded DR molecules also associated at a low level. These same species, but not DR-I chain complexes, were also shown to bind to purified HLA-DM molecules in vitro. HLA-DM interaction was quantitatively superior with DR molecules isolated in association with CLIP. DM-DR complexes generated by incubating HLA-DM with purified DR alpha beta CLIP contained virtually no associated CLIP, suggesting that this superior interaction reflects a prolonged HLA-DM association with empty class II dimers after CLIP dissociation. Incubation of peptide-free alpha beta dimers in the presence of HLA-DM was found to prolong their ability to bind subsequently added antigenic peptides. Stabilization of empty class II molecules may be an important property of HLA-DM in facilitating antigen processing.
Figures
References
-
- Unanue ER. Antigen-presenting function of the macrophage. Ann Rev Immunol. 1984;2:395–428. - PubMed
-
- Cresswell, P. 1994. Assembly, transport and function of MHC class II molecules. Annu. Rev. Immunol 12:259–293. - PubMed
-
- Jones PP, Murphy DB, Hewgill D, McDevitt HO. Detection of a common polypeptide chain in I-A and I-E sub-region immunoprecipitates. Mol Immunol. 1979;16:51–60. - PubMed
-
- Machamer CE, Cresswell P. Biosynthesis and glycosylation of the invariant chain associated with HLA-DR antigens. J Immunol. 1982;129:2564–2569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
