

Proliferation kinetics associated with T cell receptor-beta chain selection of fetal murine thymocytes
- PMID: 8976187
- PMCID: PMC2196377
- DOI: 10.1084/jem.184.6.2327
Proliferation kinetics associated with T cell receptor-beta chain selection of fetal murine thymocytes
Abstract
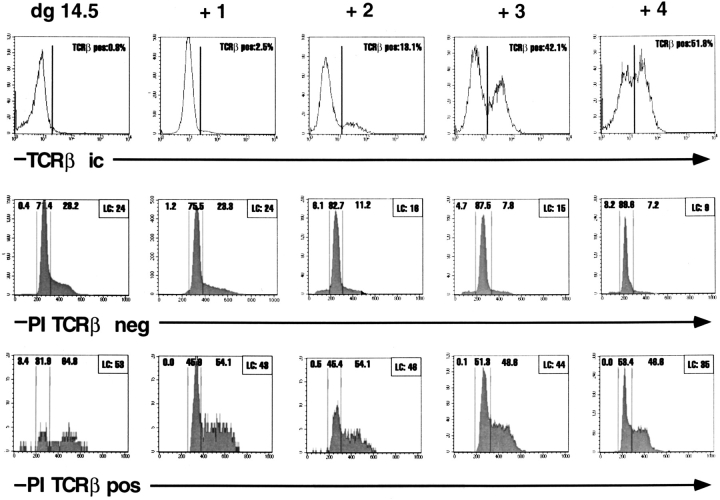
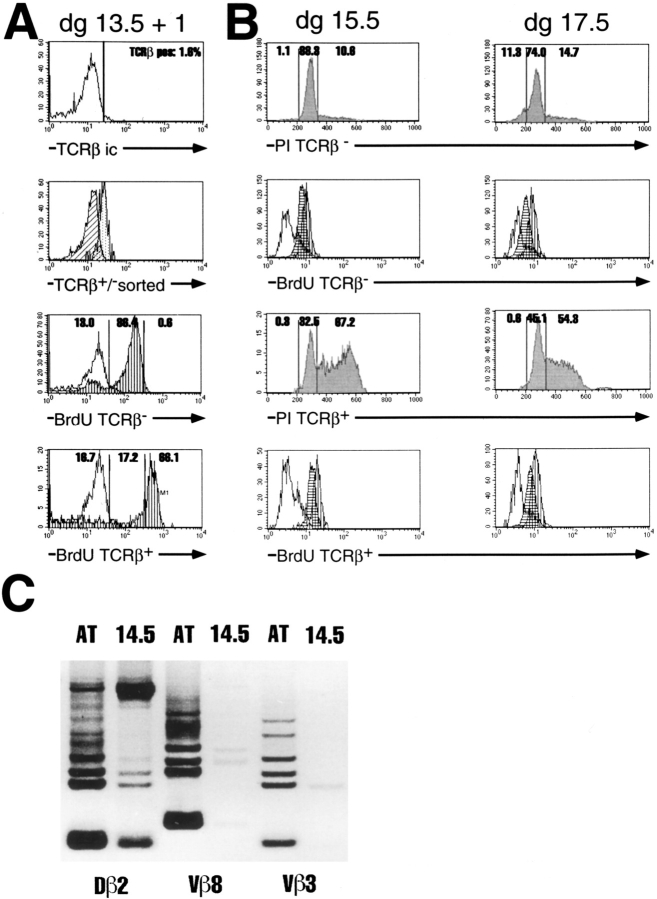
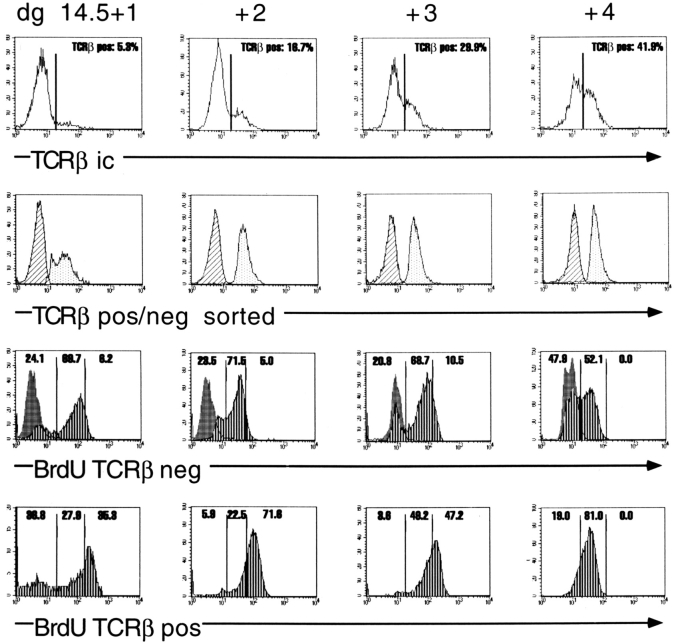
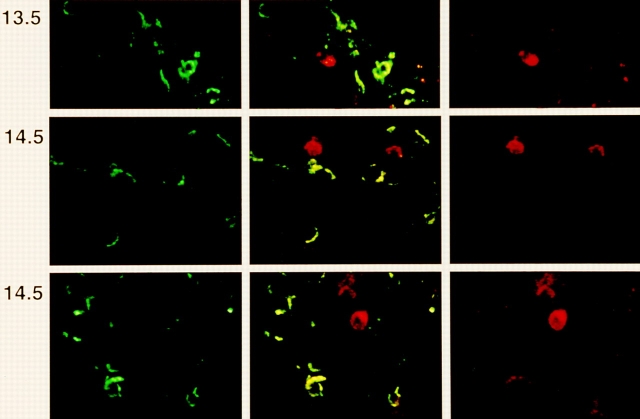
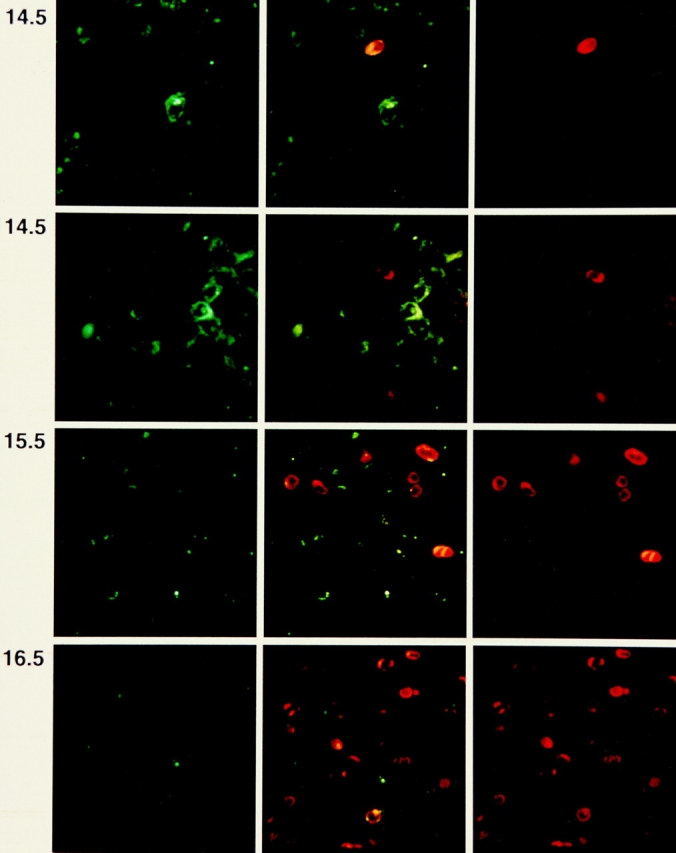
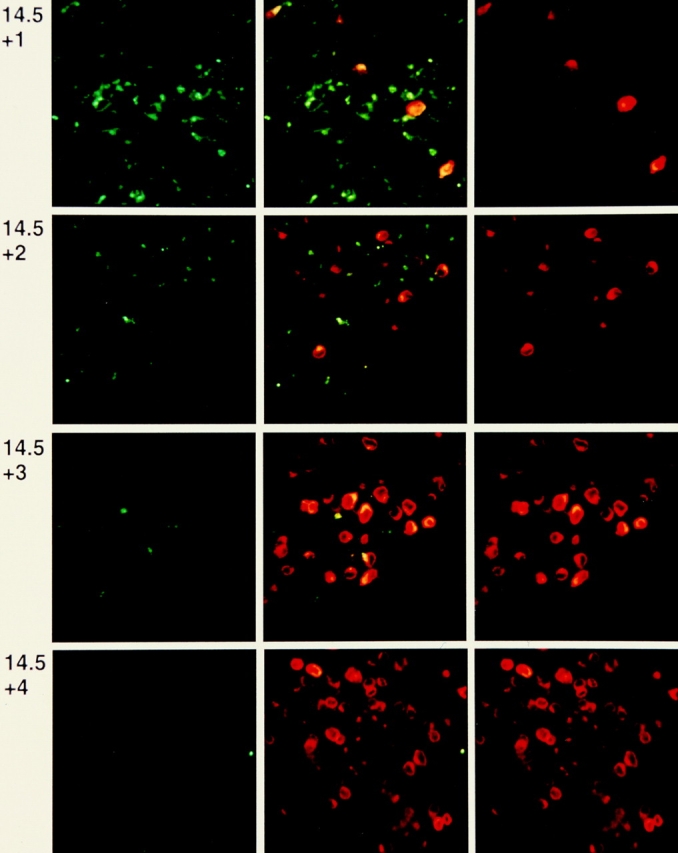
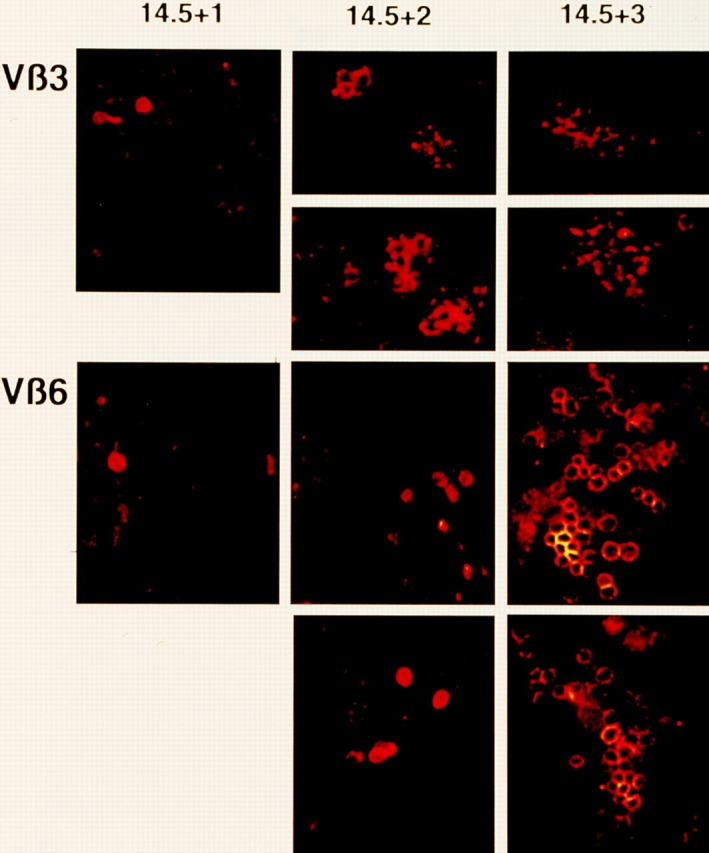
After productive rearrangement of a TCR beta chain gene, CD4-8- double negative (DN) thymocytes express TCR beta polypeptide chains on the cell surface together with pre-T alpha and the CD3 complex forming the pre-TCR. Signals transmitted through the pre-TCR select TCR beta + DN thymocytes for further maturation to the CD4+8+ double positive stage, whereas DN cells that fail to generate a productive TCR beta gene rearrangement do not continue in development. This process is termed TCR beta chain selection. Although it is likely that differences between proliferation dynamics of TCR beta + and TCR beta-cells may play a role, the exact mechanisms of TCR beta chain selection have not been elucidated. We therefore studied the proliferation dynamics of TCR beta + and TCR beta-thymocytes during fetal development, i.e., when TCR beta chain selection takes place for the first time. We analyzed in situ accumulation of TCR beta + thymocytes by confocal microscopy, and determined cell cycle and division parameters of TCR beta + and TCR beta-populations by flow cytometry. About 600 TCR beta + cells/thymic lobe are generated by independent induction events between days of gestation (dg) 13.5, and 15.5. As of dg 14.5, most TCR beta + cells have entered S/G2 phase of cell cycle, followed by seven to eight rapid cell divisions in fetal thymic organ culture, suggesting a corresponding burst of nine cell divisions within 4 d in vivo. By dg 18.5, the division rate of TCR beta + cells has slowed down to less than 1/d. About three quarters of TCR beta-cells divide at a slow rate of 1/d on dg 14.5, the proportion of nondividing cells increasing to 50% within the following four d. From dg 16.5 onwards, TCR beta-cells, but not TCR beta + cells, contain a significant proportion of apoptotic cells. The results suggest that failure to become selected results in shutdown of proliferation and eventual programmed cell death of fetal TCR beta-cells. Positive selection of fetal TCR beta + cells is achieved by an increased rate of cell divisions lasting for approximately 4 d.
Figures
References
-
- Godfrey DI, Zlotnik A. Control points in early T-cell development. Immunol Today. 1993;14:547–554. - PubMed
-
- Robey E, Fowlkes BJ. Selective events in T cell development. Ann Rev Immunol. 1994;12:675–705. - PubMed
-
- Groettrup M, Ungewiss K, Azogui O, Palacios R, Owen MJ, Hayday AC, von Boehmer H. A novel disulphide-linked heterodimer on pre–T cells consists of the T cell receptor β chain and a 33 kD glycoprotein. Cell. 1993;75:283–294. - PubMed
-
- Saint-Ruf C, Ungewiss K, Groettrup M, Bruno L, Fehling HL, von Boehmer H. Analysis and expression of a pre-T cell receptor gene. Science (Wash DC) 1994;266:1208–1212. - PubMed
-
- Groettrup M, von Boehmer H. Role for a pre–T cell receptor in T cell development. Immunol Today. 1993;14:610–614. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
