

On the molecular basis and regulation of cellular capacitative calcium entry: roles for Trp proteins
- PMID: 8986787
- PMCID: PMC26380
- DOI: 10.1073/pnas.93.26.15195
On the molecular basis and regulation of cellular capacitative calcium entry: roles for Trp proteins
Abstract
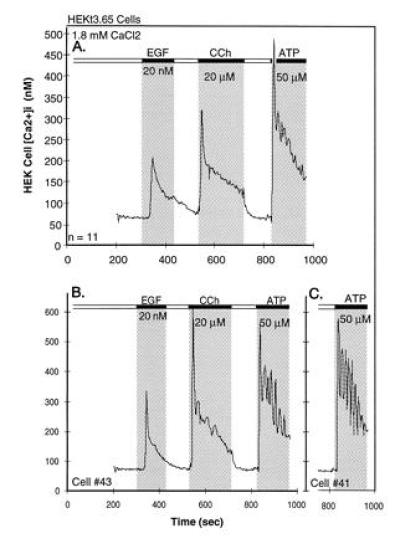
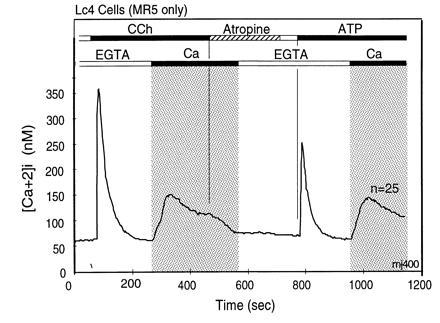
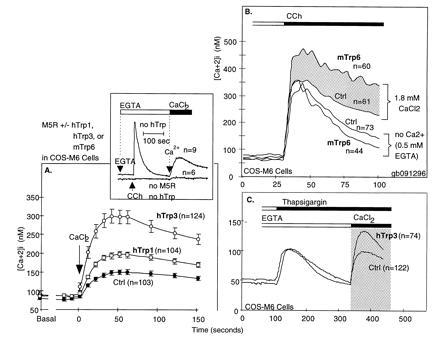
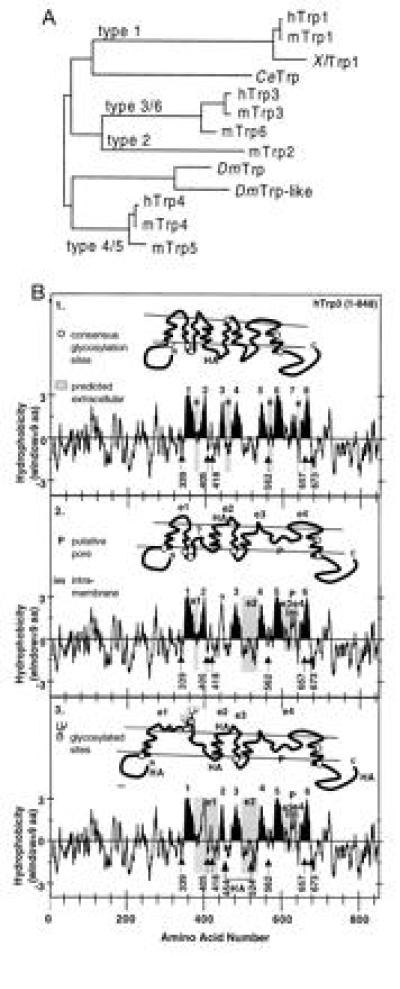
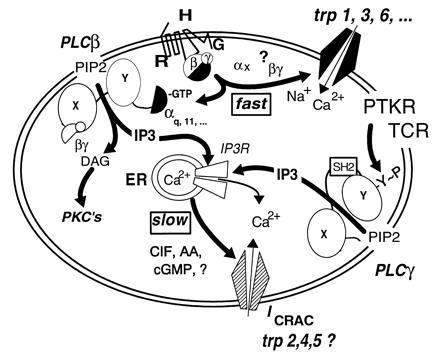
During the last 2 years, our laboratory has worked on the elucidation of the molecular basis of capacitative calcium entry (CCE) into cells. Specifically, we tested the hypothesis that CCE channels are formed of subunits encoded in genes related to the Drosophila trp gene. The first step in this pursuit was to search for mammalian trp genes. We found not one but six mammalian genes and cloned several of their cDNAs, some in their full length. As assayed in mammalian cells, overexpression of some mammalian Trps increases CCE, while expression of partial trp cDNAs in antisense orientation can interfere with endogenous CCE. These findings provided a firm connection between CCE and mammalian Trps. This article reviews the known forms of CCE and highlights unanswered questions in our understanding of intracellular Ca2+ homeostasis and the physiological roles of CCE.
Figures
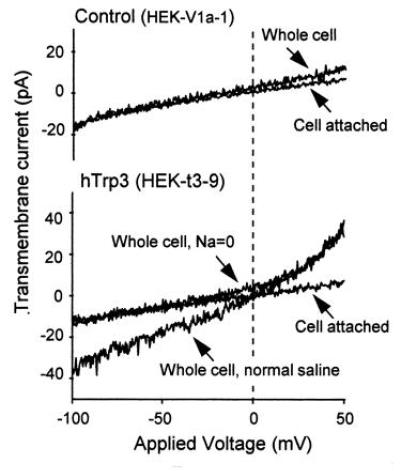
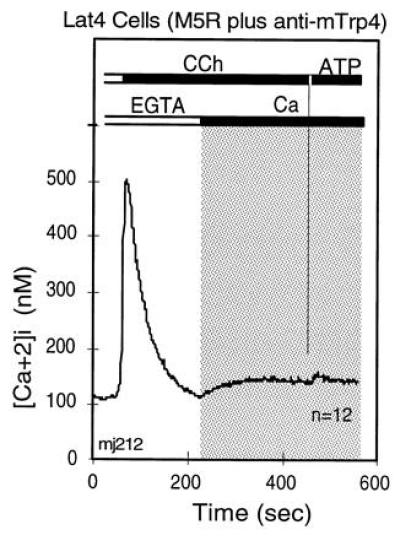
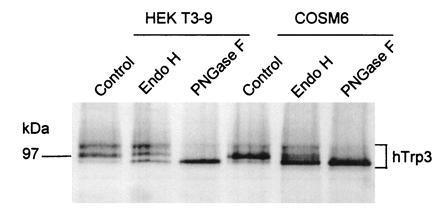
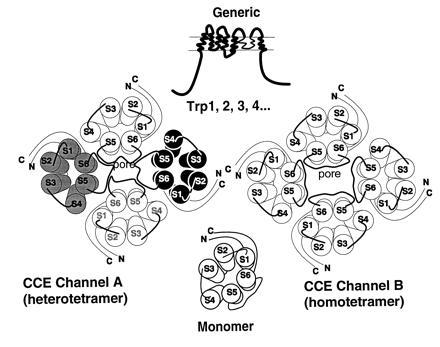
References
-
- Birnbaumer L, Campbell K P, Catterall W A, Harpold M M, Hofmann F, Horne W A, Mori Y, Schwartz A, Snutch T P, Tanabe T, Tsien R W. Neuron. 1994;13:505–506. - PubMed
-
- Perez-Reyes E, Schneider T. Kidney Int. 1995;48:1111–1124. - PubMed
-
- Seeburg P H. Trends Neurosci. 1993;16:359–365. - PubMed
-
- Fasolato C, Innocenti B, Pozzan T. Trends Pharmacol Sci. 1994;15:77–83. - PubMed
-
- Striggow F, Ehrlich B E. Curr Opin Cell Biol. 1996;8:490–495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
