

Cytotoxic T lymphocytes and viral turnover in HIV type 1 infection
- PMID: 8986810
- PMCID: PMC26403
- DOI: 10.1073/pnas.93.26.15323
Cytotoxic T lymphocytes and viral turnover in HIV type 1 infection
Abstract
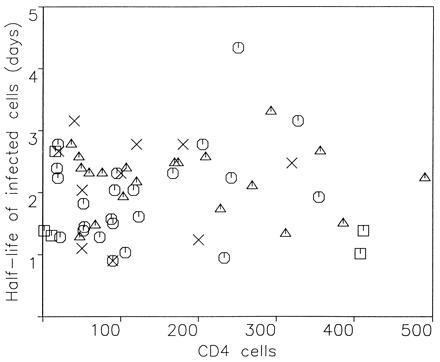
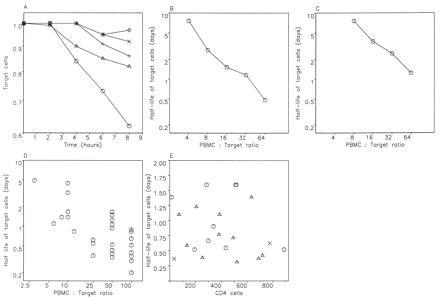
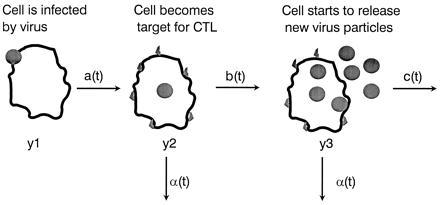
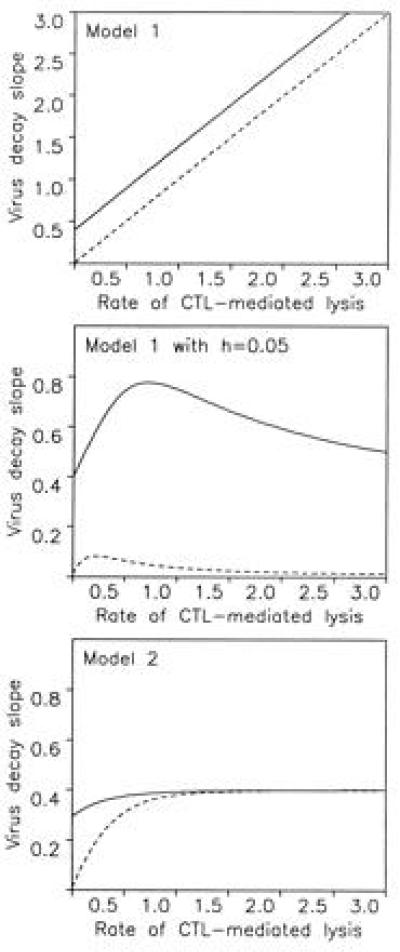
To understand the role of the immune system in limiting HIV type 1 replication, it is critical to know to what extent the rapid turnover of productively infected cells is caused by viral cytopathicity or by immune-mediated lysis. We show that uncultured peripheral blood mononuclear cells of many patients contain cytotoxic T lymphocytes (CTL) that lyse target cells-at plausible peripheral blood mononuclear cell-to-target ratios-with half-lives of less than 1 day. In 23 patients with CD4 counts ranging from 10 to 900 per microliter, the average rate of CTL-mediated lysis corresponds to a target cell half-life of 0.7 day. We develop mathematical models to calculate the turnover rate of infected cells subjected to immune-mediated lysis and viral cytopathicity and to estimate the fraction of cells that are killed by CTL as opposed to virus. The models provide new interpretations of drug treatment dynamics and explain why the observed rate of virus decline is roughly constant for different patients. We conclude that in HIV type 1 infection, CTL-mediated lysis can reduce virus load by limiting virus production, with small effects on the half-life of infected cells.
Figures
Similar articles
-
Peripheral blood cytotoxic gammadelta T lymphocytes from patients with human immunodeficiency virus type 1 infection and AIDS lyse uninfected CD4+ T cells, and their cytocidal potential correlates with viral load.J Virol. 2003 Feb;77(3):1848-55. doi: 10.1128/jvi.77.3.1848-1855.2003. J Virol. 2003. PMID: 12525619 Free PMC article.
-
Efficient lysis of human immunodeficiency virus type 1-infected cells by cytotoxic T lymphocytes.J Virol. 1996 Sep;70(9):5799-806. doi: 10.1128/JVI.70.9.5799-5806.1996. J Virol. 1996. PMID: 8709196 Free PMC article.
-
Notwithstanding Circumstantial Alibis, Cytotoxic T Cells Can Be Major Killers of HIV-1-Infected Cells.J Virol. 2016 Jul 27;90(16):7066-7083. doi: 10.1128/JVI.00306-16. Print 2016 Aug 15. J Virol. 2016. PMID: 27226367 Free PMC article.
-
What do mathematical models tell us about killing rates during HIV-1 infection?Immunol Lett. 2015 Nov;168(1):1-6. doi: 10.1016/j.imlet.2015.07.009. Epub 2015 Aug 13. Immunol Lett. 2015. PMID: 26279491 Review.
-
Perspectives on inducing efficient immune control of HIV-1 replication--a new goal for HIV therapeutics?AIDS. 2001 Feb;15 Suppl 2:S36-42. doi: 10.1097/00002030-200102002-00007. AIDS. 2001. PMID: 11424975 Review.
Cited by
-
The efficiency of the human CD8+ T cell response: how should we quantify it, what determines it, and does it matter?PLoS Comput Biol. 2012;8(2):e1002381. doi: 10.1371/journal.pcbi.1002381. Epub 2012 Feb 23. PLoS Comput Biol. 2012. PMID: 22383867 Free PMC article.
-
How is the effectiveness of immune surveillance impacted by the spatial distribution of spreading infections?Philos Trans R Soc Lond B Biol Sci. 2015 Aug 19;370(1675):20140289. doi: 10.1098/rstb.2014.0289. Philos Trans R Soc Lond B Biol Sci. 2015. PMID: 26150655 Free PMC article.
-
Prolonged activation of virus-specific CD8+T cells after acute B19 infection.PLoS Med. 2005 Dec;2(12):e343. doi: 10.1371/journal.pmed.0020343. Epub 2005 Nov 1. PLoS Med. 2005. PMID: 16253012 Free PMC article.
-
Determinants of human immunodeficiency virus type 1 escape from the primary CD8+ cytotoxic T lymphocyte response.J Exp Med. 2004 Nov 15;200(10):1243-56. doi: 10.1084/jem.20040511. J Exp Med. 2004. PMID: 15545352 Free PMC article.
-
Virus dynamics and drug therapy.Proc Natl Acad Sci U S A. 1997 Jun 24;94(13):6971-6. doi: 10.1073/pnas.94.13.6971. Proc Natl Acad Sci U S A. 1997. PMID: 9192676 Free PMC article.
References
-
- Ho D D, Neumann A U, Perelson A S, Chen W, Leonard J M, Markowitz M. Nature (London) 1995;373:123–126. - PubMed
-
- Wei X, Ghosh S K, Taylor M E, Johnson V A, Emini E A, et al. Nature (London) 1995;373:117–122. - PubMed
-
- Coffin J M. Science. 1995;267:483–489. - PubMed
-
- Nowak M A. Nature (London) 1995;375:193. - PubMed
-
- Perelson A S, Neumann A U, Markowitz M, Leonard J M, Ho D D. Science. 1996;271:1582–1586. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
